Активниот центар на протеинот и неговата интеракција со лигандот. Активниот центар на протеини и селективност на неговото врзување со лиганд
), како и во хемијата на сложените соединенија, означувајќи се таму во прилог на едно или повеќе централни (комплексни) метални атоми на честички.
Во неорганска хемија
Најчесто, таквото врзување се јавува со формирањето на таканаречената "координација" донатор-прифатената комуникација, каде што лиганди дејствуваат како основа на Луис, односно постојат донатори на електронски пар. Кога приложување на лиганди до централниот атом, хемиските својства на комплексниот агент и самите лиганд често се подложени на значителни промени.
Номенклатура ЛИГАНДОВ
- првиот во насловот на врската во номинативниот случај се нарекува анјон, а потоа и во родителскиот катјон
- во насловот на комплексот јон, лиганди први се наведени по азбучен редослед, а потоа централниот атом
- централниот атом во неутрални катјонски комплекси се нарекува руска титула, и во анјони на коренот на латинското име со суфиксот "во". По името на централниот атом, степенот на оксидација е индициран.
- бројот на лиганди прикачени на централниот атом е означен со префиксито на "моно", "ДИ", "Три", "Тетра", "Пента" итн.
Карактеристики на лиганди
Електронска структура
Всушност, најважната карактеристика Лиганд, кој овозможува да се процени и да се предвиди својата способност за сложеност и само-дисеминација на D-орбитално - уништување на соединението како целина. Во првата апроксимација, вклучува количината на електронски парови, кои лиганд можат да го распределат за создавање на координативни обврзници и електронелбилноста на дијагоналниот атом или функционална група.
Долината
Бројот на координативни места окупирани од страна на лигандот на централниот атом (или атоми) се нарекува детербинација (од LAT. дЕСТ, ДЕНТ - заб). Лиганди кои го окупираат едното координативно место се нарекуваат моноdentatat (на пример, Н.H 3), два - б.dentatat (оксалат-анјон [ О.-C (\u003d O) -C (\u003d O) -О.] 2-). Лиганди способни за окупирање големо количество Места обично укажуваат на тоа како поли.стоматолошки. На пример, етилендиаминететацетичка киселина (ЕДТА) способна за окупација на шест координативни места.
Во прилог на приемност, постои карактеристика што го одразува бројот на атоми на лиганд поврзани со една координативна локација на централниот атом. Во англиската литература е означена со зборот hapticity. и има номенклатура со соодветен индекс на натпис. Иако добро воспоставениот термин на руски јазик, очигледно нема, во некои извори можете да го најдете влечењето "Хази". Како пример, циклопентадиенил лиганд во метални централни комплекси, кои зафаќаат една координативна локација (тоа е, што е моноденти) и поврзан со сите пет јаглеродни атоми: η 5 -.
Методи на координација
 Лиганди со приемност повеќе од два се способни за формирање хелатни комплекси (Грчки. χηλή
- Cleash) - комплекси, каде централниот атом е вклучен во еден или повеќе циклуси со молекулата на лиганд. Таквите лиганди се нарекуваат хелација. Како пример, можете да ги доведете комплексите на Tetraunion на истата ЕДТА, обрнувајќи внимание на неколку од четирите m-o врски во тоа формално може да биде јонски. .
Лиганди со приемност повеќе од два се способни за формирање хелатни комплекси (Грчки. χηλή
- Cleash) - комплекси, каде централниот атом е вклучен во еден или повеќе циклуси со молекулата на лиганд. Таквите лиганди се нарекуваат хелација. Како пример, можете да ги доведете комплексите на Tetraunion на истата ЕДТА, обрнувајќи внимание на неколку од четирите m-o врски во тоа формално може да биде јонски. .
Кога се формираат хелатни комплекси често се почитува ефект на хелат - Голема стабилност во споредба со слични комплекси на лиганди кои не се хелат. Таа се постигнува поради поголемиот дел од централниот атом од замена на влијанијата и ефектот на ентропијата. На пример, дисоцијација константа на амонијак комплекс на кадмиум 2+ е речиси 1500 пати помалку од комплексот со етилендиамин 2+. Причината за ова е дека кога хидран кадмиум јонски (ii) комуницира со етилендиамин, две лигандски молекули се заменети со четири молекули на вода. Во овој случај, бројот на слободни честички во системот значително се зголемува, а ентропијата на системот се зголемува (и внатрешното нарачување на комплексот се зголемува соодветно). Тоа е, причината за хелатискиот ефект е зголемување на ентропијата на системот при замена на полидативните монотерни лиганди и, како резултат на тоа, намалената енергија на Гибс.
 Меѓу хелатни лиганди, класата на макроциклични лиганди може да се разликува - молекули со доволно за премиса комплексот на комплексот на големината на интрацикличниот простор. Пример за вакви соединенија може да послужи како порфирински бази - темелите на најважните биохемиски комплекси, како што се хемоглобин, хлорофил и бактериофил. Исто така, како макроциклични лиганди можат да се серват-етер, циклус, итн.
Меѓу хелатни лиганди, класата на макроциклични лиганди може да се разликува - молекули со доволно за премиса комплексот на комплексот на големината на интрацикличниот простор. Пример за вакви соединенија може да послужи како порфирински бази - темелите на најважните биохемиски комплекси, како што се хемоглобин, хлорофил и бактериофил. Исто така, како макроциклични лиганди можат да се серват-етер, циклус, итн.
Лиганди, исто така, може да бидат премостени, формирајќи обврзници помеѓу различни централни атоми во био или полидер комплекси. Премостените лиганди се назначени од грчкото писмо μ ( му).
Напишете преглед за статијата "Лиганд"
Белешки
|
||||||||||||||||||
Лиганд извадок
"Ако сè се бореше само со нивните убедувања, нема да има војна", рече тој."Тоа би било одлично", рече Пјер.
Принцот Андреј се насмеа.
- Тоа може многу да биде одлично, но никогаш нема да биде ...
- Па, зошто одите во војна? - го праша Пјер.
- За што? Не знам. Значи тоа е потребно. Покрај тоа, јас одам ... - на запре. - Одам, бидејќи овој живот водам тука, овој живот не е за мене!
Во соседната соба, женски фустан доби. Како да се будат, принцот Ендрју се ослободи, а неговото лице го зеде истиот израз како што имаше во дневната соба Ана Павловна. Пјер ги намали нозете од троседот. Принцезата влезе. Таа веќе беше во другата, дома, но исто како елегантен и свеж фустан. Принцот Андреј застана, разделив нејзиниот стол.
"Зошто, често мислам:" Таа зборуваше, како и секогаш, на француски, набрзина и набрзина седна на стол ", зошто не се оженил со Анет?" Како што сите сте глупави, меси, дека не се омажи за неа. Извинете, но не разбирате ништо кај жените. Каков спор сте, Monsieur Pierre.
- Јас сум со вашиот сопруг сите тврдејќи; Јас не разбирам зошто тој сака да оди во војна ", рече Пјер, без никаков срамежлив (толку обичен во односот на еден млад човек на млада жена) се свртува кон принцезата.
Принцезата беше фиксна. Очигледно, зборовите на Пјер ја допреле за живеење.
- Ох, го кажувам истото! - таа рече. - Не разбирам, не разбирам зошто човекот не може да живее без војна? Зошто ние, жените, не сакаат ништо, не ни треба ништо? Па, ти си судија. Му кажувам сè: тука е аѓутант на вујко, најбрилијантната позиција. Секој го познава толку многу, па го цениме. Пред некој ден, Aproksins, јас слушнав, како една дама прашува: "C" Est Ca Le Famex Prince Andre? " MA Brosol d "Honneur! [Дали е ова познатиот принц Андреј? Искрен збор!] - Се смееше. - Тој го прифати толку насекаде. Тоа може многу лесно да биде излез адвиерант. Знаете, Суверенот зборуваше многу милостиво со него. Разговаравме со Аненет, би било многу лесно да се организираме. Што мислиш?
Пјер го погледна принцот Андреј и, забележувајќи дека овој разговор не му се допаднал на неговиот пријател, не одговорил ништо.
- Кога ќе одиш? - тој ме праша.
- Ах! Не сум Парлез Pas de Ce Ce CE заминување, Ne M "En Reux Pas Entender Parler, [ах, не ми кажувај за ова заминување! Не сакам да слушнам за него", зборуваше принцезата во таква Каприциозно разигран тон како што зборуваше со Gippolit во дневната соба, а тоа беше толку очигледно не отиде во семејната кригла, каде што Пјер беше како член. - Денес, кога мислев дека е неопходно да се прекинат сите овие скапи односи ... и тогаш, дали знаете, Андре? - Таа значително трепери на нејзиниот сопруг. - Ј "Аи Пје, Џ" Аи Пје! [Се плашам, се плашам!] "Таа шепна, тресејќи го грбот .
Мажот ја погледна со таков вид, како да е изненаден, забележа дека некој друг, покрај него и Пјер, бил во собата; И тој со ладна учтивост прашање апелираше до неговата сопруга:
- Што се плашиш од Лиза? Не можам да разберам ", рече тој.
- Ова е како сите мажи Егоиста; Сите, сите егоисти! Тој самиот, поради неговиот каприц, Бог знае зошто ме фрла, се заклучува во селото.
"Со татко и сестра, не заборавајте", рече принцот Андреј.
- Како и да е, иако, без моите пријатели ... и тој сака да се плашам.
Нејзиниот тон веќе беше вознемирен, сунѓерот се зголеми, давајќи му на лицето не е радосно, а бруталниот, Белич, изразувајќи. Таа молчи, како да најде непристојно да зборува со Пјер за неговата бременост, тогаш како и во оваа и суштината на случајот беше.
- Сеедно, јас не го разбирам, де Quoi Vous Avez Peur, [Што се плашиш,] - полека рече принцот Андреј, не туширање на неговата сопруга.
Принцезата вцрвена и очајно ги мавташе рацете.
- Не, Андре, Џе Дис н Que vous vous Avez, промена промени ... [Не, Андреј, јас велам: ти толку промени ...]
"Вашиот лекар ви кажува да одите во кревет порано", рече принцот Андреј. - Ќе заспиете.
Принцезата не кажа ништо, и одеднаш сунѓер чевли наскоро со мустаќи; Принцот Андреј, ставајќи и отфрли, помина низ собата.
Пјер беше изненаден и наивно погледна низ очилата на него, а потоа се пресели во принцезата, како да сакаше да стане, но повторно мислеше.
"Кој е случајот што Monsiea Pierre", одеднаш рече малку принцеза, а нејзиното убаво лице одеднаш процвета во плачлива гримаса. - Ве сакав долго време, Андре: Зошто ме променивте толку многу? Што направив со вас? Одиш во армијата, не ме жалиш. За што?
- Lise! - Само рече принцот Андреј; Но, во овој збор, имаше и барање, и закана, и што е најважно, уверувањето е дека таа самата ќе одлучи според нејзините зборови; Но, таа побрза продолжи:
- Ќе се свртиш со мене, како и кај пациенти или со дете. Гледам сè. Дали некогаш сте биле пред шест месеци?
"Lise, ве замолам да престанете", рече принцот Андреј повеќе експресивен.
Пјер, се повеќе и повеќе дојдоа до возбуда за време на овој разговор, станаа и се приближуваа кон принцезата. Се чинеше дека не го толерира видот на солзите и тој беше подготвен да плаче.
- Смири, принцеза. Ви се чини дека затоа што ве уверувам, јас го доживеав ... Зошто ... затоа што ... не, жал, некој друг е излишен ... не, се смири ... Збогум ...
Принцот Андреј ја запре раката.
- Не, почекајте, Пјер. Принцезата е толку добра што тој не сака да ме лиши со задоволство да ја поминам вечерта со вас.
"Не, тој само мисли за себе", рече принцезата, без да ги држи лутите солзи.
- Лиза, - рече Духо принцот Андреј, подигање на тонот врз степенот што покажува дека трпението е исцрпено.
Одеднаш, лут регион, изразот на убава ликер на принцот беше заменет со атрактивна и возбуда на изразот на стравот; Таа ги погледна нејзините прекрасни очи на нејзиниот сопруг, а на нејзиното лице се чинеше срамежливо и препознавајќи го изразот што кучето се случува, туку слабо нервозно спуштено од опашката.
Висока специфичност на врзувачки протеин со лиганд е обезбедена со комплементарност на структурата на активниот центар на протеинската структура на лиганд
Под комплементарност, просторната и хемиската кореспонденција на интерактивните молекули разбираат. Лиганд треба да има способност да влегува и совпаѓа со конформацијата на активниот центар. Оваа случајност може да биде нецелосна, но благодарение на конформационата личност на протеинот, активниот центар е способен за мали промени и "прилагодени" под лигандот. Покрај тоа, помеѓу функционалните групи на лиганд и радикалите на амино киселини кои формираат активен центар треба да бидат поврзани со лигандот во активниот центар. Линковите помеѓу лиганд и активниот центар на протеинот можат да бидат и не-ковалентни (јонски, водород, хидрофобни) и ковалентни.
1. Карактеристики на активниот центар
Активниот центар на протеинот е релативно изолиран од околната протеинска област формирана од остатоци од аминокиселини. Во оваа област, секој остаток поради неговата индивидуална големина и функционални групи го формира "олеснувањето" на активниот центар.
Комбинацијата на такви амино киселини во еден функционален комплекс ја менува реактивноста на нивните радикали, исто како што звучните промени музички инструмент во ансамблот. Затоа, остатоците од аминокиселините вклучени во активниот центар често се нарекуваат "ансамбл" на амино киселини.
Единствените својства на активниот центар зависат не само од хемиските својства на формирањето на нејзините аминокиселини, туку и на нивната точна меѓусебна ориентација во вселената. Затоа, дури и помали нарушувања на целокупната конформација на протеинот како резултат на точка промени во неговата примарна структура или услови на животната средина може да доведе до промена на хемиските и функционалните својства на радикалите кои го формираат активниот центар, го нарушуваат врзувањето на протеините со лиганд и неговата функција. Со денатурација, активниот центар на протеините е уништен, а нивната загуба на биодиверзитетот се јавува.
Често активниот центар е формиран на таков начин што пристапот на водата на функционалните групи на неговите радикали е ограничен, односно. Условите се создаваат за да се поврзат лиганд со радикали на аминокисели.
Во некои случаи, лиганд се приклучи само на еден од атомите, што има одредена реактивност, на пример, додавање на 2 до моглобин или хемоглобинска жлезда. Сепак, својствата на овој атом селективно комуницираат со O 2 се одредуваат со својствата на радикалите околу атомот на железо во темата. ГЕМ е исто така содржан во други протеини, како што се цитохроми. Сепак, функцијата на атомот на железо во цитомомите е поинаква, таа служи како посредник за пренос на електрони од иста супстанција до друга, додека железото станува две, а потоа тривалентно.
Основната сопственост на протеините во основните функции е селективноста на прилогот кон одредени области на протеинската молекула на специфични лиганди.
2. Лиганд сорти
Лиганди може да бидат неоргански (често метали јони) и органски супстанции, ниска молекуларна тежина и високи молекуларни тежини;
постојат лиганди кои ја менуваат нивната хемиска структура при пристапот до активниот центар на протеините (промени во подлогата во активниот центар на ензимот);
постојат лиганди кои се поврзуваат со протеинот само за време на работењето (на пример, o 2, транспортирани од хемоглобин) и лиганди кои постојано се поврзани со протеини кои вршат помошна улога во функционирањето на протеините (на пример, железо, што е дел од хемоглобинот).
Во случаи кога аминокиселинските остатоци кои формираат активен центар не можат да обезбедат функционирање на овој протеин, неактивните молекули можат да бидат поврзани со одредени области на активниот центар. Така, во активниот центар на многу ензими постои јон од метал (кофактор) или органска не-зелена молекула (коензим). Не-цврст дел, цврсто поврзан со активниот центар на протеинот и неопходни за неговото работење, се нарекува "простатачка група". Миглобин, хемоглобин и цитохром имаат во активниот центар протетска група - ГЕМ со железо.
Соединението на околините во олигомерниот протеин е пример за интеракција на лиганди со висока молекуларна тежина. Секој протестен метар поврзан со други протометри служи како лиганд за нив, како и за него.
Понекогаш прилогот на било кој лиганд ја менува конформацијата на протеинот, како резултат на кој се формира обврзувачкиот центар со други лиганди. На пример, Salmodulin протеин по обврзувачки на четири јони CA 2+ во одредени сајтови, способноста да комуницирате со некои ензими, менувајќи ја нивната активност.
8. Структура на протеини. Карактеристики на структурата на функционирањето на олигомерните протеини на пример на хемоглобин. Кооперативни промени во конформацијата на демонстрантите. Можноста за регулирање на биолошката функција на олигомерните протеини со алко-цврсти лиганди.
Под кватернерната структура, начинот на поставување во просторот на поединечни полипептидни синџири, кои имаат иста (или различна) основна, секундарна или терцијарна структура и формирање на единствен во структурниот и функционален сооднос на макромолекуларното образование. Многу функционални протеини се состојат од неколку полипептински синџири поврзани со не-ковалентни обврзници, но некои валентност (слични на оние кои ја обезбедуваат стабилноста на терцијалната структура). Секој одделно се зема полипептид синџир, наречен протеер, мономер или субодит, не најчесто имаат биолошка активност. Оваа способност на протеинот стекнува со одреден метод на просторно здружение на демонстранти вклучени во неговиот состав, односно. Постои нов квалитет кој не е карактеристичен за мономерниот протеин. Како резултат на молекулата се нарекува олигомер (или мултимер). Олигомерните протеини почесто се конструираат од парен број на демонстранти (од 2 до 4, поретко од 6 до 8) со исти или различни молекуларни тежини - од неколку илјади до стотици илјади. Особено, молекулата на хемоглобинот се состои од два идентични α и два β-полипептински синџири, т.е. Тоа е тетрамер.
Кооперативни промени во конформацијата на демонстрантите.
Промена на конформацијата, а со тоа и функционалните својства на сите демонстранти на олигомерниот протеин кога лигандот е поврзан само со еден од нив, се нарекува кооперативни промени во конформацијата на протестарите.
Алостерична регулација . Ензимот ја менува активноста со помош на не-коваленлен поврзан ефект. Обврзувањето се јавува во заговор, просторно далечно од активниот (каталитичкиот) центар. Ова обврзувачки предизвикува конформациски промени во молекулата на протеини, што доведува до промена во одредена геометрија на каталитичкиот центар. Активноста може да се зголеми - ова е активирање на ензимот или намалувањето - оваа инхибиција на "пораката" на прилогот на Alto-користењето на активаторот се пренесува преку конформационите промени во каталитичката субединица, која станува комплементарна супстрат и ензимот "се врти". Кога активаторот е отстранет, ензимот се претвора во неактивна форма и "се исклучува". Алостеричната регулација е главниот метод за регулирање на метаболичките патишта.
Водородни врски, Ван дер Валл сила. Врзувањето или здружението на лиганд со рецептор (т.н. "докинг" на лиганд во специфична "ниша" во рецепторот) обично е реверзибилна и краткотрајна. Обратното процес се нарекува дисоцијација на лиганд поради рецепторот. Неразумните ковалентни врзување на лиганд со рецептор или друга молекуларна цел за овој лиганд е реткост во биолошките системи, барем во физиолошки услови. Сепак, вештачки, егзогени лиганди, неповратно ковалентно обврзувачки за целните молекули, се разбира, постојат и се дури и важни во медицината, како што е неповратно алкилирање на ДНК антитуморни лекови од алкилирајќи тип или неповратно инактивирање на Мао антидепресиви на IMAO групата, или неповратно инактивирање α - феноксибензамин адререцептори. За разлика од усвоената определба на лиганд во органометална и неорганска хемија, за интеракцијата на лиганд со целни биомолекули, апсолутно не е важно (и не е потребно), така што лиганд комуницира со металникот-кофактор во составот на биолошката Молекула (особено бидејќи не сите биолошки молекули содржат метали како кофактори). Врзувањето на лиганд е со локација која содржи метална молекула, сепак, во биолошки системи често се наоѓа и има важен биолошко значење и за транспортните протеини, како што се хемоглобин (транспорт на кислород, јаглерод диоксид и способен за транспорт на други ендогени гасови, особено ендогени јаглерод моноксид, ендогенниот водороден сулфид и ендоген сулфур оксид (IV)) и за каталитички ензими, од кои многу се металони (Содржи активен каталитички центар на јон или друг метал во координативниот комплекс со протеин).
Лиганди кои се поврзани со рецептори, но не можат или речиси можат да го активираат рецепторот (или подобро да го направат со занемарлива мала веројатност) и, соодветно, не можат да предизвикаат и не предизвикуваат физиолошкиот одговор на рецепторниот систем, туку само да ги спречат обврзувачките и двете Агонистите и инверзните агонисти и физиолошкиот одговор на нив се нарекуваат антагонисти.
Во примерот прикажан на левата страна, "ефектот на доза" зависни криви се прикажани за два лиганди со различен степен на афинитет кон рецепторот (различен афинитет кон тоа). Обврзувањето на рецепторниот лиганд често се карактеризира во смисла на која е потребна концентрација на лиганд со цел да се земат 50% од сите достапни места за врзување на рецепторите - т.н. IC 50. Вредноста на IC 50 е поврзана со констанцата за дисоцијација К I, но се разликува од него. Исто така, се разликува од вредноста на ЕЗ 50, бидејќи окупацијата од 50% од достапните рецептори не мора да води кон производство од 50% од максималниот физиолошки одговор за овој агонист, или 50% од максималниот физиолошки одговор "воопшто" (IC 50 може да биде толку поголем, така и помалку ЕЗ 50, во зависност од особеностите на регулирањето на одреден физиолошки рецепторски систем - постојат и системи за рецепторни системи во кои окупацијата на релативно мала количина на рецептори создава голем физиолошки ефект и, Напротив, треба да се преземат системи во кои треба да се преземат голем процент за да се создаде значителен физиолошки ефект. Достапните рецептори и зависноста на физиолошкиот ефект од процентот на вработување на рецепторот, како и од дозата на агонистот, не е обврзан да биде линеарен). Лиганд, кривата "ефект на доза" за која е прикажана со црвена линија, има повеќе висок степен Рефинитеции на рецепторот (поголем афинитет на врзување) од лиганд, кривата за која е прикажана со зелена линија. Ако двата лиганди се присутни во исто време, поголем процент на висока чистота (со повисок афинитет за рецепторот) лиганд ќе бидат поврзани со достапните места за врзување на рецепторите, во споредба со помалку афинитет лиганд. Овој механизам објаснува, особено зошто јаглеродниот оксид (ii) дури и при ниски концентрации може да се натпреварува со кислород за врзување на хемоглобинот, што е повисок афинитет (со поголем афинитет за хемоглобин) "агонист" на овој транспорт протеин, и зошто тоа често води на труење со јаглерод моноксид.
Афинитет на врзувањето на лиганд со рецепторот (степенот на афинитет на лиганд до рецепторот) најчесто се определува со користење на методот на поместување на етикетираниот радиоактивен лиганд (наречен "топол лиганд") на Леиниот лиганд (наречен "ладно" или "тест" лиганд). Експериментите за хомологен натпреварувачки врзувачки лиганд со рецептор се експериментите во кои "жешкото" (означено со радиоактивна етикета) и "студ" (не-етикетен) лиганд е иста хемиска супстанција, и тие се натпреваруваат едни со други за прифатлива обврзувачка сајтови со рецептор. Исто така, постојат методи без користење на радиоактивна етикета, како што се површината Пласмон резонанца, двоен поларизација интерферометрија. Овие методи овозможуваат да се утврди не само афинитетот (степенот на афинитет) на агонистот на рецепторот, туку и кинетика на нејзината асоцијација и дисоцијација поради рецепторот, а во случај на двоен поларизација интерферометрија - исто така конфигурација промени во рецептор предизвикан од врзувањето на агонистот со него. Неодамна беше развиен методот Microtermophoreseseseseses. Овој метод овозможува да се одреди афинитетот на врзувањето, без да се преклопуваат сите ограничувања на молекуларната тежина на лиганд.
Да се \u200b\u200bанализираат добиените податоци за кинетика на лиганд врз основа на рецепторот и на нејзиниот афинитет, се користат методи на статистичка механика, особено пресметката на T.N. "Конфигурација интеграл". .
Рефиниција за рецептори (афинитет) и моларна активност ("Потентност") Лиганд
Степенот на афинитет на лиганд до рецепторите, или т.н. "афинитет" на лигандот во самиот себе, сé уште не ја одредува моларната активност (општа "потентност") на еден или друг лиганд. Моларната активност (потенцијата) на супстанцијата е резултат на сложена интеракција помеѓу нејзиниот степен на афинитет кон рецепторите и нејзината внатрешна агонистичка активност (со други зборови, нејзината ефикасност на рецепторот). Внатрешната агонистичка активност (ефикасност на рецепторот) е квантитативна карактеристика на способноста на овој лиганд да предизвика еден или друг биолошки одговор по врзување на рецепторот и мерката на вредноста на биолошкиот одговор предизвикан од нив, како процент од максимум Можен биолошки одговор, за кој се зема максимална стимулација на ендогениот агонист (100%). Во зависност од природата, природата, знакот и големината на модулот предизвикан од биолошкиот одговор предизвикан од лиганд, тој е класифициран или како агонист или дури и супергонист или како делумен агонист или како неутрален антагонист или како обратен агонист или како обратен агонист или како обратен агонист или како обратен агонист или како обратен агонист или како обратен агонист или како обратен агонист или како обратен агол .
Селективни и неселективни лиганди
Селективните лиганди имаат тенденција на клинички / физиолошки релевантни (обично наномоларни) концентрации клинички / физиолошки да се обратат само прилично ограничен сет на подтипови на рецептор (не мора да бидат сите овие подтипови ќе бидат рецептори на истиот ендогенски лиганд). Во исто време, неселективните лиганди имаат тенденција во релевантни концентрации за значително да се поврзат со прилично широк спектар на подтипови на рецептори (често на различни ендогени лиганди) и, со што се произведува поширок спектар на клинички, биохемиски и физиолошки ефекти, како пожелни и, честопати несакани несакани ефекти.
Селективноста на лиганд е концепт, а условно и роднина, бидејќи има многу малку вистински селективен лиганди што се врзуваат само со еден Подтип на рецепторите во целиот спектар на "разумни", клинички остварливи концентрации кај луѓето, па дури и помалку лиганди способни за одржување на 100% селективност во оние концентрации кои можат да се креираат во експериментите со животни и особено "во тест цевка" ( in vitro.). Често очигледна релативна селективност на еден или друг лиганд се губи кога се зголемува дозата или концентрацијата (односно во повисоки концентрации или дози, почнува да комуницира со други подтипови на рецептори) и има важна клиничка вредност (така, Високите дози на селективната аспект на опиоидните рецептори на бупренорфин се способни за тоа дека е значително лути на дишењето и предизвика еуфорија, бидејќи селективноста е изгубена во споредба со морфинот; на сличен начин, високи дози на селективни β-adrenobloclockers се способни за Обезбедување на бронхоспазам, бидејќи селективноста е изгубена од подтипот β 1, а високите дози на β 2 -годиеностимуланти во прилог на елиминирање на бронхоспазмот, исто така, може да предизвика тахикардија; високи дози на атипични антипсихотици како што се рисперидон и оланзапин се способни да предизвикаат екстрапирамидни несакани ефекти, како типични антипсихотици).
Развојот на нови, повеќе селективни, лиганди е важна задача на модерната експериментална и клиничка фармакологија, бидејќи селективните лиганди, селективно активирање или блокирање на само еден "неопходен" подтип на рецептори или неколку од нивните подтипови, имаат тенденција да покажат помалку несакани ефекти, додека Неселективни лиганди, обврзувачки со широк спектар на рецептори, произведуваат и пожелни и несакани несакани ефекти. Добар пример Тоа е споредба со релативно неселективна хлорпромазин со повеќе селективен халоперидол: хлорпромазин, поради неговата ниска селективност, произведува многу несакани ефекти во прилог на корисен антипсихотичен ефект (така, α 1-Adrenoblocade води до хипотензија и тахикардија, H1-Histamic Blockade за поспаност, седација, зголемување на апетитот и зголемување на тежината на телото, M-cholinyblocade - за да се исуши устата и запек, итн., Додека халоперидол, овие феномени значително предизвикуваат клинички употребувани дози, главно екстрапирамидни несакани ефекти директно поврзани со неговата главна дејство од 2-блокирање).
Мерката на релативната селективност на овој или оној лиганд е вредноста на нејзиниот афинитет (афинитет) на "посакуваниот", "главен" подтип на рецептори (на пример, до D 2, во случај на антипсихотици), и на Најблискиот по редослед на афинитет на афинитетот на афинитетот (афинитет) подтип рецепторите - тоа е, вредноста на односот k i (1) / k i (2). Повисоки филиси на "посакуваниот" тип на рецептори, повеќе високо активни ("повисоки прецизност") соединенија често, иако не секогаш, се повеќе селективни, барем во ниски концентрации (употребата на која, повторно, станува можно благодарам на повеќе од висок афинитет соединение во однос на рецепторот и поголема активност за поврзување). Така, важната задача на експериментална и клиничка фармакологија е развојот на новиот, повисок Phyphine (поседува повисок афинитет за рецепторот) и поактивни ("повеќе споменати") во однос на тие или други видови на рецептори, соединенија.
Бивалентни лиганди
Бивалентни лиганди се состојат од две поврзани молекули, од кои секоја е лиганд за одреден подтип на рецептори (исти или различни), и врз основа на карактеристиките на просторна структура, двата дела на молекулата се способни за во исто време се врзуваат на два дела од "композитен" хомо или хетеродимички рецептор комплекс. Бивалентни лиганди се користат во научни истражувања со цел да се открие и да ги проучат рецепторите хомо-и хетеродимерски комплекси и да ги проучуваат нивните својства. Бивалентни лиганди обично се големи молекули и имаат тенденција да имаат својства потребни за лекови, како што се погодно фармакокинетиката (прифатлива биорасположивост, погодност на клиничката употреба, прифатлив полуживот, итн.), Ниска алергичност и прифатлива токсичност и несакани ефекти кои Ги прави, како по правило, несоодветни или несоодветни за употреба во клиничка пракса, надвор од истражувачките лаборатории.
Привилегирана структура
Преферираната структура е структурниот дел од молекулата, радикалниот или хемиски елементкој е статистички повторен меѓу веќе познатите лекови на оваа фармаколошка класа, меѓу веќе познатите лиганди од овој тип или подтип на рецептори или познати инхибитори на овој ензим, или меѓу некои други освен некој вид општи знаци Специфичен подмножество на веќе познати биолошки активни соединенија. Овие статистички изолирани привилегирани елементи на хемиската структура може дополнително да се користат како основа за развој на нови биолошки активни соединенија или нови лекови со слични или, можеби, подобрени својства, па дури и да се развијат цели библиотеки на таквите соединенија.
Карактеристични примери се, на пример, трицикличните структури од различна хемиска структура во составот на трицикличните молекули на трицикличните антидепресиви или постоењето на хемиски слични интеграции на антипсихотиците, како што се деривати на Бутизон (Hanoperidol, Spireton, droperidol, итн (Резервни, карбидин, итн.), Деривати на фенотиазинот (хлорпромазин, перфеназин итн.).
исто така види
Белешки
- Teif v.b. (2005). "Ligand-индуцирана ДНК кондензација: изборот на моделот". Биофизички весник. 89 (4): 2574-2587. DOI: 10.1529 / Biophysj.105.063909. PMC. PMID.
- Teif VB, Rippe K. (2010). "Модели за статистички механички решетки за вршење на протеини-ДНК во хроматин". Весник на физиката: кондензирана материја. 22 (41): 414105.
Модул структура. | Теми |
Модуларна единица 1. | 1.1. Структурна организација на протеини. Фази на формирање на мајчин протеински конформации 1.2. Основи на протеини. Дрога како лиганди кои влијаат на функцијата на протеини 1.3. Денатурација на протеини и можноста за нивната спонтана ренативација |
Модуларна единица 2. | 1.4. Карактеристики на структурата и функционирањето на олигомерните протеини на пример на хемоглобин 1.5. Одржување на мајчин конформација на протеините во клеточните услови 1.6. Различни протеини. Семејство на протеини на пример на имуноглобулини 1.7. Физичко-хемиски својства на протеини и методи на нивно одвојување |
Модуларна единица 1 Структурна организација на мономерни протеини и основи на нивното функционирање
Целите на студијата се способни да:
1. Користете познавања за карактеристиките на структурата на протеините и зависноста на функциите на протеините од нивната структура за да ги разберете развојните механизми на наследни и стекнати протеинопатии.
2. Објаснете ги механизмите за терапевтско дејство на некои лекови како лиганди кои комуницираат со протеини и разликување на нивната активност.
3. Користете знаење за структурата и конформационата личност на протеините за да ја разберат нивната структурна и функционална нестабилност и тенденција за денатурација во променливите услови.
4. Проширување на употребата на агенти за денатурирање како средства за стерилизирање на медицински материјал и алатки, како и антисептици.
Знам:
1. Нивоа на структурната организација на протеини.
2. Вредноста на примарната структура на протеините кои го дефинираат нивниот структурен и функционален колектор.
3. Механизмот на формирање во протеините на активниот центар и неговата специфична интеракција со лигандот што го потценува функционирањето на протеините.
4. Примери за ефектот на егзогени лиганди (лекови, токсини, отрови) за конформацијата и функционалната активност на протеините.
5. Причини и последици од протеинските денатури, фактори за денатурација.
6. Примери за користење на фактори за денари во медицината како антисептици и средства за стерилизирање на медицински инструменти.
Тема 1.1. Структурна организација на протеини. Фази на мајчин формирање
Конформации на протеините
Протеините се полимерни молекули чии мономери се само 20 α-амино киселини. Поставувањето и редот на соединението на амино киселини во протеинот се одредува со структурата на гените во ДНК на поединци. Секој протеин во согласност со неговата специфична структура ја врши карактеристиката карактеристична за тоа. Комплетните протеини на овој организам ги одредуваат неговите фенотипски карактеристики, како и присуството на наследни болести или предиспозиција за нивниот развој.
1. Амино киселини вклучени во протеините. Пептид врска.Протеини - Полимери изградени од мономери - 20 α-амино киселини, општа формула кои
Амино киселините се разликуваат во структурата, големини, физичко-хемиски својства на радикалите прикачени на α-јаглеродниот атом. Функционалните групи на амино киселини ги одредуваат карактеристиките на својствата на различни α-амино киселини. Радикалите пронајдени во α-амино киселини можат да се поделат во неколку групи:
Пролин.за разлика од другите 19 мономери на протеини, а не амино киселина, но имино киселина, радикал во пролин е поврзан и со α-јаглероден атом и со Imino Group
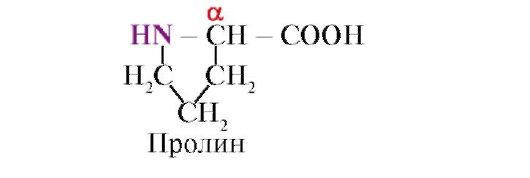 Амино киселините се разликуваат во растворливоста во вода.Ова се должи на способноста на радикалите да комуницираат со вода (хидрат).
Амино киселините се разликуваат во растворливоста во вода.Ова се должи на способноста на радикалите да комуницираат со вода (хидрат).
ДО хидрофилнирадикалите кои содржат анјонски, катјонски и поларни функционални групи.
ДО хидрофобрадикали кои содржат метил групи, алифатички синџири или циклуси.
2. Пептидните обврзници ги поврзуваат амино киселините во пептиди.Во синтезата на пептид, α-карбоксилна група од една амино киселина комуницира со α-амино група на друга амино киселина за да се формира пептидна комуникација:
Протеините се полипептиди, т.е. Линеарни полимери на α-амино киселини поврзани со пептидна врска (Слика 1.1.)
 Сл. 1.1. Термини кои се користат во опишувањето на структурата на пептидите
Сл. 1.1. Термини кои се користат во опишувањето на структурата на пептидите
Се нарекуваат амино киселински мономери вклучени во полипептидите амино киселини остатоци.Синџир на повторувачки групи - NH-CH-CO- Форми пептид јадро.Амино киселински остатоци со бесплатна α-амино група се нарекува N-терминал, и има бесплатна α-карбоксилна група - C-терминал. Пептидите се снимаат и читаат од N-Terminus до C-END.
Пептидната врска формирана од Пролинмино групата се разликува од другите пептидни обврзници: не постои водород во азотниот атом на пептидната група,
наместо тоа, има врска со радикал, како резултат на тоа, едната страна од циклусот е вклучена во пептидниот кабел:
 Пептидите се разликуваат од амино киселински состав, количината на амино киселини и редоследот на соединението на амино киселини, на пример, серма-де-ГИС и ГИС-ГИС-Ала-Сери-Сери се два различни пептиди.
Пептидите се разликуваат од амино киселински состав, количината на амино киселини и редоследот на соединението на амино киселини, на пример, серма-де-ГИС и ГИС-ГИС-Ала-Сери-Сери се два различни пептиди.
Пептидните врски се многу издржливи, а строгите услови се потребни за нивната хемиска nefermental хидролиза: анализираниот протеин е хидролизиран во концентрирана хлороводородна киселина на температура од околу 110 ° 24 часа. Во жива ќелија, пептидните врски може да се растурат со протеолитички ензимиповикан протеаниили пептидхидролази.
3. Примарна протеинска структура.Аминокиселините остатоци во пептидните синџири на различни протеини не се случајно, но се наоѓаат во одреден редослед. Се нарекува линеарна секвенца или редот на алтернација на остатоци од аминокиселини во полипептидниот синџир примарна протеинска структура.
Примарната структура на секој поединечен протеин е кодирана во молекулата на ДНК (во парцела наречена ген) и се спроведува за време на транскрипцијата (препишување на информации за MRNA) и превод (синтеза на примарна протеинска структура). Следствено, примарна структура на поединците на поединецот - информации за утврдување на карактеристиките на структурата на протеините на ова тело, што зависи од функцијата на постоечките протеини од родителите до децата, што зависи од функцијата на протеините ( Слика 1.2.).
 Сл. 1.2. Односот помеѓу генотипот и конформацијата на протеините синтетизирани во телото на поединецот
Сл. 1.2. Односот помеѓу генотипот и конформацијата на протеините синтетизирани во телото на поединецот
Секој од околу 100.000 индивидуални протеини во човечкото тело има уникатенпримарна структура. Во молекулите на еден вид на протеини (на пример, албумин), истиот албум на остатоци од аминокиселини, кој го разликува албуминот од кој било друг индивидуален протеин.
Секвенцата на остатоци од аминокиселини во пептидниот синџир може да се гледа како форма на снимање на информации. Оваа информација го одредува просторно поставување на линеарен пептид синџир во покомпактна три-димензионална структура наречена конформацијаверверица. Се нарекува процес на формирање функционално активна конформација на протеинот преклопување.
4. Конформација на протеини.Слободната ротација во пептидната изолација е можна помеѓу азотниот атом на пептидната група и соседниот атом на α-јаглерод, како и помеѓу α-јаглеродниот атом и јаглерод карбонил група. Поради интеракцијата на функционалните групи на остатоци од аминокиселини, примарната структура на протеините може да се здобијат со посложени просторни структури. Во глобуларни протеини се разликуваат две главни нивоа на поставување на конформација на пептидните синџири: среднои. терцијарни структури.
Секундарна структура на протеини- Ова е просторна структура која е формирана како резултат на формирањето на водород обврзници помеѓу функционалните групи -C \u003d O и - NH-пептид. Во овој случај, синџирот на пептид може да стекне редовни структури од два вида: α-спиралаи. β-структури.
Внатре α-спиралаводородните обврзници се формираат помеѓу атомот на кислород на карбонилната група и водородот на азот азот 4 од амино киселината; Странични синџири на остатоци од аминокиселини
тие се наоѓаат по периферијата на спиралата, без да учествуваат во формирањето на секундарната структура (слика 1.3.).
Волуметриски радикали или радикали кои носат идентични трошоци спречуваат формирање на α-хеликс. Остатокот од пролин со прстен структура ја прекинува α-спиралата, бидејќи поради недостаток на водород на азотниот атом во пептидниот синџир е невозможно да се формира водород обврзница. Односот помеѓу азот и атомот на α-јаглерод е дел од циклусот на пролин, така што пептидот на јадрото на ова место станува виткање.
β-структуратаа е формирана помеѓу линеарните региони на островот Пептид на еден полипептид синџир, формирајќи преклопени структури. Полипептински синџири или делови може да се формираат паралелноили анти-паралелни β-структури.Во првиот случај, N- и C-краевите на дистрибуираните пептидни синџири се совпаѓаат, а во вториот - тие имаат спротивна насока (Слика 1.4).
Сл. 1.3. Средна протеинска структура - α-спирала
 Сл. 1.4. Паралелни и анти-паралелни β-преклопени структури
Сл. 1.4. Паралелни и анти-паралелни β-преклопени структури
β-структури се означени со широк стрелки: A - анти-паралелна β-структура. Б - паралелни β-преклопени структури
Во некои, протеините на β-структурата можат да се формираат со формирање на водород обврзници помеѓу атомите на пептидниот остров со различни полипептидни синџири.
Во протеините се наоѓаат исто така области со неправилно средноструктурата на која се свиоци, јамки, врти на островот Полипептид. Тие често се наоѓаат на места каде што насоката на пептидните синџир промени, на пример, при формирање паралелна β-преклопена структура.
Според присуството на α-хеликс и β-структури, глобуларните протеини може да се поделат во четири категории.
Сл. 1.5. Секундарната структура на миоглобинот (а) и β-синџирот на хемоглобин (б) кои содржат осум α-спирали

 Сл. 1.6. Секундарна структура на трисофосфатизомераза и пируваткиза домен
Сл. 1.6. Секундарна структура на трисофосфатизомераза и пируваткиза домен
 Сл. 1.7. Секундарна структура на постојаниот домен на имуноглобулин (а) и ензимскиот суперксидазмаза (б)
Сл. 1.7. Секундарна структура на постојаниот домен на имуноглобулин (а) и ензимскиот суперксидазмаза (б)
Внатре четврта категоријавклучени протеини кои имаат незначителен број на редовни секундарни структури во нивниот состав. Таквите протеини вклучуваат мали, богати со цистеински протеини или металопротеини.
Терцијарна протеинска структура- типот на конформација, формиран од интеракциите помеѓу радикалите на амино киселини, кој може да биде на значително растојание едни од други во синџирот на пептид. Повеќето протеини во исто време формираат просторна структура слична на глобул (глобуларни протеини).
Бидејќи радикалите на хидрофобните амино киселини имаат тенденција да се комбинираат со помош на т.н. хидрофобни интеракциии интермолекуларни ван дер Валс сили, густо хидрофобно јадро се формира во внатрешноста на протеинскиот глобул. Хидрофиличните јонизирани и не-јонизирани радикали главно се наоѓаат на површината на протеинот и ја одредуваат својата растворливост во вода.
 Сл. 1.8. Видови на врски кои произлегуваат помеѓу амино киселински радикали во формирањето на терциерната протеинска структура
Сл. 1.8. Видови на врски кои произлегуваат помеѓу амино киселински радикали во формирањето на терциерната протеинска структура
1 - јонска комуникација- се јавува помеѓу позитивно и негативно наелектризирани функционални групи;
2 - водороден комуникации- се појавува помеѓу хидрофилната непоколеблива и друга хидрофилна група;
3 - хидрофобни интеракции- се јавуваат помеѓу хидрофобни радикали;
4 - дисулфид комуникации- формирана со оксидација на sh-групи на цистеински остатоци и нивна интеракција едни со други
Хидрофилни аминокиселини остатоци кои се најде во внатрешноста на хидрофобниот кернел може да комуницираат едни со други со јонски.и. водородните врски(Слика 1.8).
Јонските и водородни обврзници, како и хидрофобни интеракции се однесуваат на бројот на слаби: нивната енергија малку ја надминува енергијата на термичкото движење на молекулите на собна температура. Конформацијата на протеинот е поддржана од појавата на многу вакви слаби врски. Бидејќи атомите од кои се содржат протеини постојано движењеМожно е да се скршат слабите обврзници и формирање на други, што доведува до мало движење на поединечни делови од полипептидниот синџир. Овој имот на протеини ја менува сообразноста како резултат на кршењето на некои и се нарекува формирање на други слаби врски конформациона личност.
Во човечкото тело, системите работат на поддршка хомеостаза- Постојаност на внатрешниот медиум во одредени граници кои се дозволени за здрав организам. Под условите на хомеостазата, малите промени во конформацијата не ја нарушуваат целокупната структура и функцијата на протеините. Се нарекува функционална активна конформација на протеини мајчин конформација.Промена на внатрешниот медиум (на пример, концентрации на гликоза, јони, протони, итн.) Води кон промена во конформацијата и оштетените функции на протеините.
Терциерна структура на некои протеини се стабилизира дисулфид обврзнициформирана од интеракцијата на -шо групи од два остатоци
 Сл. 1.9. Формирање на дисулфидна врска во молекулата на протеини
Сл. 1.9. Формирање на дисулфидна врска во молекулата на протеини
цистеин (слика 1.9). Повеќето интрацелуларни протеини немаат во терцијарна структура на ковалентни дисулфид обврзници. Присуството е карактеристично за протеините што ги избираа клетките, кои ја обезбедуваат нивната поголема стабилност во екстрацелуларните услови. Така, дисулфидните обврзници се достапни во инсулин и имуноглобулински молекули.
Инсулин- Протеински хормон, синтетизиран во β-клетките на панкреасот и се секретира во крвта како одговор на зголемувањето на концентрацијата на гликоза во крвта. Во инсулинската структура, постојат две дисулфидни обврзници кои ги поврзуваат полипептидот A- и B-синџири и една дисулфидна врска во рамките на А-синџирот (слика 1.10).
 Сл. 1.10. Дисулфид обврзници во инсулин структура
Сл. 1.10. Дисулфид обврзници во инсулин структура
5. Структура на протеини од надзор.Во различни примарни структури и функции, понекогаш се откриваат протеините слични комбинации и толкување на секундарните структури,кои се нарекуваат супер-предупредувачка структура. Таа зафаќа средна позиција помеѓу секундарните и терцијарните структури, бидејќи ова е специфична комбинација на елементи на секундарната структура во формирањето на терциерната протеинска структура. Изворови структури имаат специфични имиња, како што се "α-спирала-ротирање-а-спирала", "леуцински патент", "цинк прсти", итн. Таквите супер-функционални структури се карактеристични за протеините на ДНК-обврзувачки.
"Леуцински патент".Овој тип на супер-стандардна структура се користи за поврзување на два протеини. На површината на интерактивните протеини постојат α-спирални делови кои содржат најмалку четири остатоци од леуцин. Остатоците од леуцин во α-спирали се наоѓаат во шест амино киселини еден од другиот. Бидејќи секоја α-хеликс серпентина содржи 3,6 амино киселини остатоци, радикалите на леуцин се на површината на секој втор пресврт. Остатоците од леуцин од α-хеликс од еден протеин можат да комуницираат со остатоци од леуцин на други протеини (хидрофобни интеракции), поврзувајќи ги заедно (слика 1.11.). Многу врзани протеини на ДНК функционираат како дел од олигомерните комплекси, каде што индивидуалните субоди се поврзани со едни со други "леуцин Клас".
 Сл. 1.11. "Леуцински патент" помеѓу α-спиралните делови на два протеини
Сл. 1.11. "Леуцински патент" помеѓу α-спиралните делови на два протеини
Пример за такви протеини можат да служат како хистони. Хистонс- нуклеарни протеини кои вклучуваат голем број позитивно наелектризирани амино киселини - аргинин и лизин (до 80%). Хистонските молекули се комбинираат во олигомерни комплекси кои содржат осум мономери кои користат "леуцински прицврстувачи", и покрај значајното полнење на овие молекули.
"Цинк прст"- варијанта на супер-стандардната структура карактеристика на протеините за врзување на ДНК-врзувања има форма на издолжен фрагмент на површината на протеините и содржи околу 20 амино киселини (слика 1.12). Формата на "издолжениот прст" одржува атом на цинк поврзан со радикалите на четири амино киселини - два од остатоците од цистеин и два хистидин. Во некои случаи, наместо остатоците на хистидин се остатоците од цистеин. Двајца тесно лежечки цистеински остатоци се одделени од два други остатоци од хицилили цикоселитивноста која се состои од околу 12 аминокиселини остатоци. Овој дел од протеинот формира α-спирала, чија радикали може специјално да се поврзе со регулаторните делови на големиот жлеб на ДНК. Приватни обврзувачки специфичност
 Сл. 1.12. Примарната структура на делницата на ДНК-обврзувачки протеини што го формираат структурата на "цинк прст" (буквите се означени со амино киселини вклучени во оваа структура)
Сл. 1.12. Примарната структура на делницата на ДНК-обврзувачки протеини што го формираат структурата на "цинк прст" (буквите се означени со амино киселини вклучени во оваа структура)
регулаторниот ДНК обврзувачки протеин зависи од секвенцата на остатоци од аминокиселини лоцирани во областа на "цинк прстот". Таквите структури содржат, особено, рецепторите на стероидните хормони кои се вклучени во регулативата за транскрипција (читање информации од ДНК на РНК).
Тема 1.2. Основи на протеини. Дрога како лиганди кои влијаат на функцијата на протеини
1. Активниот центар на протеинот и неговата интеракција со лигандот.Во процесот на формирање на терциерна структура на површината на функционално активниот протеин, тоа е обично во проценката, заговор формиран од страна на амино киселински радикали, далеку од едни со други во примарната структура. Овој заговор кој има единствена структура за овој протеин и способен за специфични комуницира со специфична молекула или група слични молекули се нарекува центар за врзување на протеини со лиганд или активен центар. Лиганди се нарекуваат молекули кои комуницираат со протеини.
Висока специфичностинтеракцијата на протеините со лиганд е обезбедена со комплементарност на структурата на активниот центар на структурата на лиганд.
Комплементација- Тоа е просторна и хемиска усогласеност на површините за интеракција. Активниот центар не само што треба да се поврзе со лиганд вклучен во него, туку и помеѓу функционалните групи радикали вклучени во активниот центар, а лиганд мора да биде формиран со комуникација (јонски, водород, како и хидрофобни интеракции) кои го задржуваат лиганд во активниот центар (Слика 1.13).
 Сл. 1.13. Комплементарна интеракција на протеини со лиганд
Сл. 1.13. Комплементарна интеракција на протеини со лиганд
Некои лиганди, приклучувајќи се кон активниот центар на протеините, вршат помошна улога во функционирањето на протеините. Таквите лиганди се нарекуваат кофактори и протеини кои имаат не-невообичаен дел - комплексни протеини(За разлика од едноставните протеини кои се состојат само од протеини). Не се нарекува не-истрошен дел, цврсто поврзан со протеини, се нарекува протетска група.На пример, составот на миоглобинот, хемоглобинот и цитомомите содржат цврсто прикачен на активниот центар протетска група - скапоцен камен кој содржи железо јон. Комплексните протеини кои содржат дијаманти се нарекуваат хемопротеини.
Кога е поврзан со протеини на специфични лиганди, функцијата на овие протеини се манифестира. Значи, албуминот е најважниот плазма протеин - ја манифестира својата транспортна функција, поврзувајќи ги хидрофобните лиганди во активниот центар, како што се масни киселини, билирубин, некои лекови итн. (Слика 1.14)
Лиганди во интеракција со тридимензионалната структура на пептидниот синџир не само што може да биде ниска молекуларна тежина органски и неоргански молекули, туку и макромолекули:
ДНК (примери кои се сметаат погоре со ДНК-обврзувачки протеини);
Полисахариди;
 Сл. 1.14. Интерконекција на генотипот и фенотипот
Сл. 1.14. Интерконекција на генотипот и фенотипот
Единствената примарна структура на човечките протеини кодирани во молекулата на ДНК во клетките се реализира во форма на уникатна конформација, структурата на активниот центар и функциите на протеините
Во овие случаи, протеинот препознава одредена површина на лиганд, комбиниран и комплементарен обврзувачки центар. Така, на површината на хепатоцитите, постојат протеини рецептори на хормонски инсулин, кој исто така има протеинска структура. Интеракцијата на инсулинот со рецепторот предизвикува промена во нејзината конформација и активирање на сигналните системи што доведуваат до поштенски марки во нутритроцити на хранливи материи по оброците.
На овој начин, основата на функционирањето на протеините е специфичната интеракција на активниот центар на протеинот со лиганд.
2. Структура на доменот и неговата улога во функционирањето на протеините.Долгите полипептидни синџири на глобуларни протеини често се преклопуваат во неколку компактни, релативно независни области. Тие имаат независна терцијарна структура слична на такви во глобуларни протеини и се нарекуваат домени.Благодарение на доменската структура на протеините, нивната терцијарна структура е полесна.
Во доменските протеини, обврзувачките центри со лиган често се наоѓаат помеѓу домените. Така, трипсин е протеолитичен ензим кој е произведен од егзокрин дел од панкреасот и е неопходен за варење на протеините на храната. Таа има дводимензионална структура, а Центарот за врзување на Tripsin со неговиот лиганд - протеини во храна - се наоѓа во жлебот помеѓу двата домени. Активниот центар ги создава потребните услови за ефективно врзување на специфичен дел од протеинот на храна и хидролиза на нејзините пептидни врски.
Различни домени во протеини при интеракција на активниот центар со лиганд може да се преместат во однос на едни со други (Слика 1.15).
Хексокиназ- ензим катализирање на глукоза фосфорилација користејќи АТП. Активниот центар на ензимот се наоѓа во расцепувањето помеѓу двата домени. Кога врзувачки хексидохинази со гликоза, околните домени се затворени и подлогата се појавува како "стапица", каде што се јавува фосфорилација (види слика 1.15).
 Сл. 1.15. Врзување на домени на хексидохинази со гликоза
Сл. 1.15. Врзување на домени на хексидохинази со гликоза
Некои домени протеини вршат независни функции, обврзувачки за разни лиганди. Таквите протеини се нарекуваат мултифункционални.
3. Лекови - лиганди кои влијаат на функцијата на протеини.Интеракцијата на протеините со лиганди е конкретно. Меѓутоа, поради конформационата личност на протеинот и неговиот активен центар, можно е да се избере друга супстанција која исто така може да комуницира со протеинот во активниот центар или друг дел од молекулата.
Супстанцијата, според структурата слична на природниот лиганд, се нарекува структурен аналог на лигандаили не-лиганд. Исто така, комуницира со протеинот во активниот центар. Структурен аналог на лиганд може да се подобри функцијата на протеини (агонист),затоа, намалете ја (антагонист).Лиганд и нејзините структурни аналози се натпреваруваат едни со други за врзување на протеини во еден центар. Таквите супстанции се нарекуваат конкурентни модулатори(регулатори) Protein функции. Многумина лекови дејствува како протеински инхибитори. Некои од нив се добиени со хемиска модификација на природни лиганди. Инхибитори на протеински функции може да бидат лекови и отрови.
Атропин е конкурентен инхибитор на М-холинорецептори.Acetylcholine е невротрансмитер на преносот на нервен пулс преку холинергични синапси. За да се спроведе возбуда, ацетилхолинот избран за синаптичкиот слот треба да комуницира со протеинот - постсинуптичен мембрански рецептор. Беа пронајдени два вида cholinoreCeptors:
М-рецептор,во прилог на ацетилхолинот селективно во интеракција со мускарин (токсин токсин на мукумот). М - холинорецептори се достапни на мазни мускули и кога интеракција со ацетилхолин ги тера да ги намалат;
N-рецептор,конкретно обврзувачки со никотин. H-cholinoreceptors се наоѓаат во синапс на попречни скелетни мускули.
Специфичен инхибитор М-холинорецепторие атропин. Таа е содржана во растенијата убав и бел.
 Атропин има во структурата слична на функционалните групи на ацетилхолинот и нивната просторна локација, затоа се однесува на конкурентните инхибитори на М-холинорецептор. Со оглед на тоа што врзувањето на ацетилхолинот со М-холинорецептори предизвикува намалување на мазни мускули, атропинот користи како лек кој го ублажува нивниот спазам (антиспазмодика).Значи, познато е дека користи атропин за да ги релаксира окото на очите при гледањето на ДНК на очите, како и да се отстранат спазмите со стомак колика. М-холинорецептори се исто така во централниот нервен систем (ЦНС), затоа големи дози на атропин може да предизвикаат несакана реакција од ЦНС: моторни и ментални возбудувања, халуцинации, напади.
Атропин има во структурата слична на функционалните групи на ацетилхолинот и нивната просторна локација, затоа се однесува на конкурентните инхибитори на М-холинорецептор. Со оглед на тоа што врзувањето на ацетилхолинот со М-холинорецептори предизвикува намалување на мазни мускули, атропинот користи како лек кој го ублажува нивниот спазам (антиспазмодика).Значи, познато е дека користи атропин за да ги релаксира окото на очите при гледањето на ДНК на очите, како и да се отстранат спазмите со стомак колика. М-холинорецептори се исто така во централниот нервен систем (ЦНС), затоа големи дози на атропин може да предизвикаат несакана реакција од ЦНС: моторни и ментални возбудувања, халуцинации, напади.
Ditylin е конкурентен антиколинорецептор агонист инхибиција на функцијата на невромускулните синапси.
 Нервните мускулни синапси на скелетните мускули содржат n-холинорецептори. Нивната интеракција со ацетилхолин води кон контракции на мускулите. Во некои хируршки операции, и ендоскопските студии користат лекови кои предизвикуваат релаксација на скелетни мускули (Миосонта).Тие вклучуваат DITHILINE, што е структурен аналог на ацетилхолин. Таа е придружено од N-холинорецептори, но, за разлика од ацетилхолинот, многу полека е уништена од ензимот - ацетилхолинезаза. Како резултат на долгото отворање на јонски канали и деполаризација на решетката на мембраната, нервниот импулс е нарушен и се јавува мускулната релаксација. Првично, овие својства беа откриени во отровна корара, така што таквите лекови се јавуваат одземени.
Нервните мускулни синапси на скелетните мускули содржат n-холинорецептори. Нивната интеракција со ацетилхолин води кон контракции на мускулите. Во некои хируршки операции, и ендоскопските студии користат лекови кои предизвикуваат релаксација на скелетни мускули (Миосонта).Тие вклучуваат DITHILINE, што е структурен аналог на ацетилхолин. Таа е придружено од N-холинорецептори, но, за разлика од ацетилхолинот, многу полека е уништена од ензимот - ацетилхолинезаза. Како резултат на долгото отворање на јонски канали и деполаризација на решетката на мембраната, нервниот импулс е нарушен и се јавува мускулната релаксација. Првично, овие својства беа откриени во отровна корара, така што таквите лекови се јавуваат одземени.
Тема 1.3. Денатурација на протеини и можноста за нивната спонтана ренативација
1. Бидејќи мајчин конформација на протеините се одржува поради слабите интеракции, промената во составот и својствата на животната средина на медиумот, ефектите од хемиските реагенси и физичките фактори предизвикуваат промена во нивната конформација (имотот на конформационата моќност). Прекинувањето на голем број врски води до уништување на мајчин конформација и денатурација на протеините.
Денатурација на протеините- Ова е уништување на нивната мајчин конформација под дејство на агентите за денатури предизвикани од јаз на слаби врски кои ја стабилизираат просторна структура на протеинот. Денатурацијата е придружена со уништување на уникатната три-димензионална структура и активниот центар на протеинот и губењето на нејзината биолошка активност (Слика 1.16).
Сите денатуриран молекули на протеини стекнуваат случајна конформација, која се разликува од другите молекули на истиот протеин. Амино киселински радикали кои формираат активен центар се отстрануваат едни од други, т.е. Специфичен протеински обврзувачки центар со лиганд е уништен. Со денатурација, примарната структура на протеините останува непроменета.
Употребата на агенти за денатурирање во биолошките истражувања и медицината.Во биохемиските студии, пред определувањето на соединенија со ниска молекуларна тежина, протеините обично се отстрануваат од растворот. За таа цел најчесто се користи трихлороцетна киселина (TCH). По додавањето на TCH на растворот, денатурираните протеини спаѓаат во талог и лесно се отстрануваат со филтрација (Табела 1.1.)
Во медицината, агентите за денатурирање често се користат за стерилизирање на медицинскиот инструмент и материјал во автоклави (агент за денатурирање - висока температура) и како антисептици (алкохол, фенол, хлор) за обработка на контаминирани површини кои содржат патогени микрофлора.
2. Спонтана протеинска бубрежна- Доказ за определување на примарната структура, усогласеност и функција на протеини. Индивидуалните протеини се производи од еден ген, кои имаат идентична аминокиселинска секвенца и во ќелијата ја стекнуваат истата конформација. Основен заклучок дека во примарната структура на протеинот веќе поставил информации за неговата конформација и функции, таа била направена врз основа на способноста на некои протеини (особено, рибонуклеаз и миоглобин) на спонтана ренативација - реставрацијата на нивниот роден конформација по денатурацијата.
Формирањето на просторни протеини структури се врши со методот на само-собрание - спонтан процес, во кој полипептидниот синџир кој има единствена примарна структура има за цел да усвои конформација со најмалата конформација слободна енергија. Способноста да се намалат протеините кои ја зачувуваат примарната структура по денатурацијата е опишана во експериментот со ензимската рибонуклеаза.
Рибонуклеа е ензим кој го уништува односот помеѓу индивидуалните нуклеотиди во молекулата на РНК. Овој глобуларен протеин има еден полипептид синџир, чија терцијарна структура се стабилизира со мноштво слаби и четири дисулфид обврзници.
Третманот на рибонуклеаза уреа, уништување на водород обврзници во молекулата и намалување на агентот кој ги солли дисулфидните обврзници води до денатурација на ензимот и губење на нејзината активност.
Отстранувањето на агентите за денатурирање на дијализа води кон реставрација на конформацијата и функцијата на протеини, т.е. на ренастирањето. (Слика 1.17).
 Сл. 1.17. Денатурација и повредени рибонуклеаза
Сл. 1.17. Денатурација и повредени рибонуклеаза
А - мајчин конформација на рибонукlease, во терцијарна структура на која има четири дисулфидни обврзници; Б - денатурирана молекула за рибонуклет;
V - Менативната молекула за рибонуклет со обновена структура и функција
1. Пополнете Табела 1.2.
Табела 1.2. Класификација на амино киселини на поларитетот на радикалите
2. Напишете формула на тетрапептид:
ASP - Pro - Fen-Liz
а) Означете ги групите за повторување на пептидот што го формираат пептидното јадро и променливите групи претставени со радикали на амино киселини;
б) назначете N- и C-завршува;
в) нагласи пептидните обврзници;
г) напише друг пептид кој се состои од истите амино киселини;
г) брои го бројот можни опции Тетрапептид со сличен состав на аминокиселината.
3. Објаснете ја улогата на примарната структура на протеините за примерот на компаративна анализа на две слични во структурата и еволуциони блиски пептидни хормони на неврохипофизата на цицачијата - окситоцин и вазопресин (Табела 1.3).
Табела 1.3. Структура и функции на окситоцин и вазопресин
 За ова:
За ова:
а) споредете го составот и редоследот на амино киселините од два пептиди;
б) да најдат сличности на примарната структура на две пептиди и сличноста на нивната биолошка акција;
в) да ги пронајде разликите во структурата на два пептиди и разликата помеѓу нивните функции;
г) Земете го заклучокот за ефектот на примарната структура на пептидите на нивните функции.
4. Опишете ги главните фази на формирањето на конформацијата на глобуларните протеини (секундарна, терцијарна структура, концептот на супер-стандардната структура). Наведете ги видовите на врски вклучени во формирањето на протеински структури. Кои радикали на амино киселини можат да учествуваат во формирањето на хидрофобни интеракции, јонски, водородни обврзници.
Дајте примери.
5. Дајте дефиниција на "конформационата личност на протеините", наведете ги причините за неговото постоење и вредност.
6. Проширување на значењето на следната фраза: "Функционирањето на протеините се заснова на нивната специфична интеракција со лиганд", користејќи ги условите и објаснувајќи ја нивната вредност: протеинска конформација, активен центар, лиганд, комплементарност, протеинска функција.
7. Во еден пример, објасни што се домени и каква е нивната улога во функционирањето на протеините.
Задачи за самоконтрола
1. Постави натпревар.
Функционална група во амино киселински радикал:
А. карбоксил група Б. Хидроксил група во Гуанидиноваја група на Тиолнија Група Д. Аминог
2. Изберете ги вистинските одговори.
Амино киселини со поларни непознати радикали - ова е:
А. ЦИС Б. АСН
Б. ГЛ. Три
3. Изберете ги вистинските одговори.
Амино киселински радикали:
A. Обезбедување на специфичност на примарната структура Б. Учествувајте во формирањето на терцијарната структура
Б. Се наоѓа на површината на протеинот, влијае на нејзината растворливост на G. формираат активен центар
Д. учествува во формирањето на пептидните врски
4. Изберете ги вистинските одговори.
Хидрофобни интеракции може да се формираат помеѓу амино киселински радикали:
A. Tre Lei B. Про три
Б. Се сретна со Иле Г. ТИР АЛА Д. Шакција на вратило
5. Изберете ги вистинските одговори.
Јонските врски може да се формираат помеѓу амино киселински радикали:
А. GLN ASP Б. АПР ЛИЗ
Б. Лиз г-дин ГИС АСП Д. АПР АПР
6. Изберете ги вистинските одговори.
Водородните обврзници може да се формираат помеѓу амино киселински радикали:
A. SERM GLN B. CIS TRE
Б. ASP Liz G. G-ASP D. ASN TRE
7. Постави натпревар.
Тип на комуникација вклучен во формирањето на протеинска структура:
А примарна структура Б. секундарна структура
Б. Терцијарна структура
Г. Структура на надзор на надзор Д. Конформација.
1. Водородните врски помеѓу атомите на островот Пептид
2. Слаби врски помеѓу функционалните групи на амино киселински радикали
3. Односи помеѓу α-амино и α-карбоксилни групи на амино киселини
8. Изберете ги вистинските одговори. Tripsin:
А. Протендичен ензим Б. содржи два домени
Б. Хидролилат скроб
Активниот центар се наоѓа помеѓу домените. Д. се состои од два полипептидни синџири.
9. Изберете ги вистинските одговори. Атропин:
A. NeuroTiator.
Б. Структурен аналог на ацетилхолин
Б. Интеракција со n-холинорецептори
Г. Го зајакнува нервниот пулс преку холинергичните сини
D. Конкурентен M-HOLNORECEPTOR инхибитор
10. Изберете ги вистинските изјави. Во протеини:
A. Примарна структура содржи информации за структурата на активниот центар
Б. Активниот центар е формиран на ниво на примарна структура
Б. Конформација е строго фиксирана од ковалентни обврзници
G. Active Center може да комуницира со група слични лиганди
поради конформационата побарување на протеините D. Промена на животната средина, може да влијае на афинитетот на активни
центар до Лиганда
1. 1-б, 2-Г, 3-б.
3. А, Б, Б, Г.
7. 1-б, 2-D, 3-а.
8. А, Б, Б, Г.
Главни термини и концепти
1. Протеин, полипептид, амино киселини
2. Примарна, секундарна, терцијарна протеинска структура
3. Конформација, мајчин протеински конформации
4. Ковалентни и слаби врски во протеини
5. Конформаторна Личност
6. Центар за активни верверица
7. Лиганди
8. Преклопување на Белков
9. Структурни аналози на лиганди
10. Домен протеини
11. Едноставни и комплексни протеини
12. денатурација на протеини, агенти за денатурирање
13. Реитизација на протеини
Решавање на задачите
"Структурната организација на протеини и темелите на нивното функционирање"
1. Главната функција на протеинот е хемоглобин А (NVA) - превоз на кислород во ткивата. Во популацијата на луѓето, повеќе форми на овој протеин со изменети својства и функција се познати - т.н. абнормални хемоглобини. На пример, е утврдено дека хемоглобинот, откриен во еритроцитите на пациенти со анемија со српска клетка (HBS), има ниска растворливост под услови на низок притисок на кислород (како што се одвива во венска крв). Ова води кон формирање на агрегати на овој протеин. Протеинот ја губи својата функција, паѓа во талог, а еритроцитите стекнуваат погрешна форма (некои од нив го формираат обликот на српот) и побрзо од вообичаените уништени во слезината. Како резултат на тоа, се развива анемија на Crespovid Cell.
Единствената разлика во примарната структура на NVA и откриена во делот N-терминал на β-синџирот на хемоглобин. Споредба на N-терминалните делови на β-синџирот и покажат како промените во примарната протеинска структура влијаат врз неговите својства и функции.
 За ова:
За ова:
а) Напишете ги формулите за амино киселини, според кои NVA се разликуваат и ги споредуваат својствата на овие амино киселини (поларитетот, полнење).
б) Земи го заклучокот за причината за намалување на растворливоста и нарушувањето на кислородниот транспорт во ткивото.
2. Сликата ја покажува шемата на структурата на протеинот кој има обврзувачки центар со лиганд (активен центар). Објаснете зошто протеинот има селективност во изборот на лиганд. За ова:
а) се сеќавам што е активниот центар на протеинот и ја разгледува структурата на активниот центар на протеинот презентирани на сликата;
б) да напише радикални формули на амино киселини кои се дел од активниот центар;
в) Нацртајте лиганд кој конкретно би можел да комуницира со активниот центар на протеинот. Наведете ги функционалните групи кои можат да формираат врски со амино киселински радикали кои се дел од активниот центар;
г) ги наведува видовите на врски кои произлегуваат помеѓу лиганд и радикалите на амино киселините на активниот центар;
д) Објаснете што е базирана специфичноста на интеракцијата на протеините со лиганд.
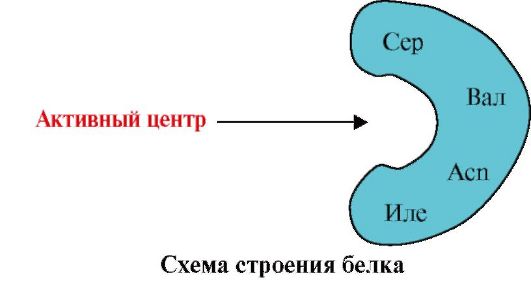 3.
Сликата го покажува активниот центар на протеините и неколку лиганди.
3.
Сликата го покажува активниот центар на протеините и неколку лиганди.
Одреди кој од лиганди со најголема веројатност ќе комуницира со активниот протеински центар и зошто.
 Кои видови врски се јавуваат во процесот на формирање на протеински лигандски комплекс "?
Кои видови врски се јавуваат во процесот на формирање на протеински лигандски комплекс "?
4. Структурните аналози на природни лиганди на протеини може да се користат како лекови за промена на активноста на протеините.
Acetylcholine е медијатор на пренесување на возбуда во невромускулни синапси. Во интеракцијата на ацетилхолин со протеини - рецепторите на постсинаптичната мембрана на скелетни мускули, отворањето на јонските канали и мускулната контракција се јавува. Ditylin е лек кој се користи во некои операции за релаксирање на мускулите, бидејќи го нарушува преносот на нервниот пулс преку невромускулни синапси. Објаснете го механизмот на дејство на dithiline како дрога miorexing. За ова:
а) напишете ги формулите на ацетилхолин и дитинилин и споредете ги нивните структури;
б) Опишете го механизмот на релаксирачко дејство на дитилин.
5. Во некои болести, пациентот ја зголемува температурата на телото, која се смета за заштитна реакција на телото. Сепак, високите температури се штетни за протеините на организмот. Објаснете зошто на температура над 40 ° C, функцијата на протеини е нарушена и се јавува заканата за човечкиот живот. За да го направите ова, запомнете:
1) структурата на протеините и комуникацијата, држејќи ја својата структура во мајчин конформација;
2) Како се менува структурата и функцијата на протеините кога е подигната температурата?;
3) Што е Хоместаза и зошто е важно да се задржи човековото здравје.
Модуларна единица 2 Олигомерни протеини како цел на регулаторни влијанија. Структурна и функционална разновидност на протеини. Методи на одвојување и чистење на протеини
Целите на студијата се способни да:
1. Користете знаење за карактеристиките на структурата и функциите на олигомерните протеини за да ги разберете адаптивните механизми за регулирање на нивните функции.
2. Објаснете ја улогата на ѕидовите во синтезата и одржувањето на конформацијата на протеините во клеточните услови.
3. Објаснете го разновидноста на манифестацијата на разновидноста на структурите и функциите што се синтетирајќи во телото на протеините.
4. Анализирајте го поврзувањето на структурата на протеините со нивната функција на примерите на споредувањето поврзани со хемопротеини - миглобин и хемоглобин, како и претставници на петте класи на протеини на семејството на имуноглобулин.
5. Примени ги знаењата за особеностите на физичко-хемиските својства на протеините за да ги изберете методите за чистење од други протеини и нечистотии.
6. Интерпретирање на резултатите од квантитативниот и квалитативниот состав на крвните плазма протеини за да ја потврди или разјасни клиничката дијагноза.
Знам:
1. Карактеристики на структурата на олигомерните протеини и адаптивни механизми за регулирање на нивните функции на примерот на хемоглобинот.
2. Структурата и функциите на хапериите и нивната вредност за одржување на мајчин конформација на протеините во клеточните услови.
3. Принципите на комбинирање на протеините во семејството за да се слични на нивната усогласеност и функции на примерот на имуноглобулините.
4. Методи на поделба на протеините врз основа на карактеристиките на нивните физичко-хемиски својства.
5. Крвна плазма електрофореза како метод за проценка на квалитативниот и квантитативниот состав на протеините.
Тема 1.4. Карактеристики на структурата и функционирањето на олигомерните протеини на пример на хемоглобин
1. Многу протеини имаат неколку полипептински синџири во нивниот состав. Таквите протеини се нарекуваат олигомери индивидуални синџири - демонстранти.Protteers во олигомерни протеини се поврзани со множество на слаби не-вирулентни обврзници (хидрофобни, јонски, водород). Интеракција
протомерите се изведуваат благодарам комплементарностнивните контактни површини.
Бројот на прототели во олигомерни протеини може да варира во голема мера: хемоглобинот содржи 4 протопромет, аспартаминотрансфераза ензим е 12 протоометриски, а 2120 протераници поврзани со не-членови врски се вклучени во тутунскиот мозаичен протеин протеин. Како резултат на тоа, олигомерните протеини може да имаат многу голема молекуларна тежина.
Интеракцијата на еден протеер со другите може да се гледа како посебен случај на интеракција помеѓу протеинот со лиганд, бидејќи секој мирис служи како лиганд за други прототели. Износот и начинот на поврзување на протометарот во протеинот се нарекуваат кватернерна протеинска структура.
Протеините може да ги вклучуваат истите или различни заштитници, на пример, хомодими - протеини кои содржат два идентични протерувачи и хетеродими - протеини кои содржат два различни протерувачи.
Ако протеините вклучуваат различни прототели, тогаш тие можат да се разликуваат во структурата на обврзувачки центри со различни лиганди. Кога врзуваат лиганди со активен центар, функцијата на овој протеин се манифестира. Центарот, кој се наоѓа на друг демонстрант, се нарекува алохектички (други различни од активни). Обврзувачки S. алостеричен лиганд или ефектор,таа врши регулаторна функција (Слика 1.18). Интеракцијата на алостеричниот центар со ефектот предизвикува конформациски промени во структурата на целиот олигомерниот протеин поради неговата конформациона дозвола. Ова влијае на афинитетот на активниот центар на специфичен лиганд и ја регулира функцијата на овој протеин. Промената во конформацијата и функциите на сите заштитни лица во интеракцијата на олигомерниот протеин барем со еден лиганд е името на кооперативните промени во конформацијата. Се нарекуваат оператори за зајакнување на протеините активатории ефектите кои ја депресираат функцијата - инхибитори.
Така, олигомерните протеини, како и протеините кои имаат структура на доменот, имотот се појавува нови во споредба со мономерните протеини - способноста за алтогретерска регулација на функциите (регулирање со приложување кон протеинот на различни лиганди). Ова може да се следи со споредување на структурите и функциите на два блиски сложени протеини на миоглобинот и хемоглобинот.
 Сл. 1.18. Структура на димензински протеини
Сл. 1.18. Структура на димензински протеини
2. Формирање на просторни структури и функционирање на миоглобинот.
Миглобин (МВ) - протеини лоцирани во црвени мускули, чија главна функција е создавање на резерви од 2 потребни со интензивна мускулна работа. МВ е сложен протеин кој го содржи протеинскиот дел - Апом и не-пилешки дел - ГЕМ. Примарната структура на АПМУ ја одредува својата компактна глобуларна конформација и структурата на активниот центар на кој се приклучи не-квалификуваниот дел од миглобинот. Кислород кој доаѓа од крв во мускулите е обврзувачки за FE + 2 Gema како дел од миоглобинот. МВ е мономерно протеин кој има многу висок афинитет за O 2, според тоа, враќањето на кислородот од страна на миоглобинот се јавува само со интензивна мускулна работа, кога делумниот притисок o 2 се намалува нагло.
Формирање на MV конформација.Во црвените мускули на рибозомите, за време на емитувањето, синтезата на примарната структура на МВ, претставена со специфичен редослед од 153 аминокиселини остатоци. Средното структура на МВ содржи осум α-спирали, наречени латински букви од А до Н, помеѓу кои постојат некаризирани делови. Терциерна структура на МВ има форма на компактен глобул, во продлабочувањето на кој се наоѓа активен центар помеѓу F и E α-хеликс (слика 1.19).
 Сл. 1.19. Mioglobin структура.
Сл. 1.19. Mioglobin структура.
3. Карактеристики на структурата и работењето на активниот центар на МВ.МВ активниот центар е претежно хидрофобни аминокиселински радикали, далеку едни од други во примарната структура (на пример, три 3 9 двете сушари за коса 138) до активниот центар се споени со слабо растворливи во вода лиганди - дијамант и околу 2. ГЕМ е специфичен лиганд на АПМУ (Слика 1.20), чија основа е четири пиролни прстени поврзани со метрички мостови; Во центарот постои атом на Fe + 2, поврзан со атомите на азот на пиролеските прстени со четири координативни обврзници. Во активниот центар на МВ, покрај радикалите на хидрофобните амино киселини, постојат и остатоци од две амино киселини со хидрофилни радикали - ГИС Е 7.(ГИС 64) и ГИС Ф 8.(ГИС 93) (Слика 1.21).
 Сл. 1.20. Структурата на GEM-не-откриениот миглобин и хемоглобин
Сл. 1.20. Структурата на GEM-не-откриениот миглобин и хемоглобин
 Сл. 1.21. GEMA и O 2 локација во активниот центар на апомоглобин и хемоглобин
Сл. 1.21. GEMA и O 2 локација во активниот центар на апомоглобин и хемоглобин
ГЕМ преку атом на железо е ковалентно поврзан со ГИС Ф 8. O 2 се приклучува на жлездата на другата страна на авионот на Хем. ГИС Е 7 е неопходно за правилна ориентација од 2 и го олеснува додавањето на кислород до Fe + 2 полите
ГИС Ф 8.форми Координација со FE + 2 и цврсто го поправа GEM во активниот центар. ГИС Е 7.ние сме неопходни за соодветна ориентација во активниот центар на друг лиганд - O 2 кога ќе комуницира со Fe + 2 Gema. GEMA микро-операција создава услови за издржлив, но реверзибилен обврзувачки o 2 со Fe +2 и ја спречува водата во хидрофобниот активен центар, што може да доведе до негово оксидација во FE + 3.
Структурата на мономер на МВ и нејзиниот активен центар го одредува високиот афинитет на протеинот до 2.
4. Олигомерната структура на HB и регулирање на афинитетот на HB до 2 лиганди. Човечки хемоглобини- семејство на протеини, како и миглобин поврзани со комплексни протеини (хемопротеини). Тие имаат структура на тетрамер и содржат два α-синџири, но се разликуваат во структурата на два други полипептидни синџири (2α-, 2x синџири). Структурата на вториот полипептиден синџир ги одредува карактеристиките на функционирањето на овие форми на HV. Околу 98% од хемоглобинот на еритроцитите на возрасни е хемоглобин А.(2α-, 2P синџири).
За време на периодот на интраутерина развој, функционираат два главни вида хемоглобини: ембрионски NV.(2α, 2ε), кој се наоѓа во раните фази на развојот на фетусот, и хемоглобин f (фетал)- (2α, 2γ), кој доаѓа да го замени почетокот на хемоглобинот на фетусот на шестиот месец на интраутерина развој и само по раѓањето е заменето со NV A.
NV A - протеин, поврзан миглобин (MV), е содржан во еритроцитите на возрасни. Структурата на нејзините индивидуални протери е слична на онаа на миглобинот. Средното и терцијарната структура на миоглобините и покровитерите на хемоглобините се многу слични, и покрај фактот што само 24 амино киселини се идентични во примарната структура на нивните полипептидни синџири (секундарната структура на демонстрантите на хемоглобинот, како и миглобинот, содржат осум α- Спирали означени со латински букви од п.н.е. и терцијарна структура има форма на компактен глобул). Но, за разлика од миоглобинот, хемоглобинот има олигомерна структура, се состои од четири полипептински синџири поврзани со не-ковалентни обврзници (слика 1.22).
Секој Protiber HB е поврзан со не-откриен дел од Gem и соседните демонстранти. Согорувањето на протеинскиот дел на HV со дијамант е сличен на оној на миглобинот: во активниот центар на протеинот, хидрофобните ГЕМ хидрофобни делови се опкружени со хидрофобни амино киселини со исклучок на ГИС Ф 8 и ГИС Е 7 , кои се наоѓаат на двете страни на авионот на Хем и играат слична улога во функционирањето на протеинот и врзување со кислород (види структура на миоглобинот).
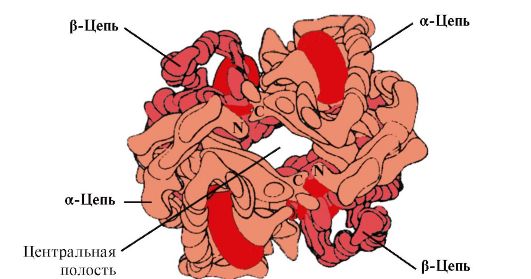 Сл. 1.22. Олигомерна структура на хемоглобин
Сл. 1.22. Олигомерна структура на хемоглобин
Згора на тоа, ГИС Е 7.врши важни дополнителна улогаво функционирањето на HB. Слободен дијамант има 25.000 пати повисок афинитет за CO од K O 2. Ко во мали количини се формира во телото и, со оглед на својот висок афинитет за Г., тој може да го прекрши превозот на потребните клетки за животот на 2. Сепак, како дел од афинитетот на хемоглобинот на полите на јаглеродниот оксид го надминува афинитетот од 2 само 200 пати поради присуството во активниот центар на ГИС Е 7. Остатокот од оваа амино киселина создава оптимални услови Да се \u200b\u200bповрзе со Heme со O 2 и ја ослабува интеракцијата на HEME со CO.
5. Главната функција на НВ е превоз околу 2 од белите дробови во ткаенината.За разлика од мономерниот миоглобин, со многу висок афинитет кон O 2 и ја врши функцијата на кислород во црвени мускули, олигомерната структура на хемоглобинот обезбедува:
1) брза сатурација на HV кислород во белите дробови;
2) способноста на HB да даде кислород во ткивата на релативно висок парцијален притисок o 2 (20-40 mm Hg. Art.);
3) можност за регулирање на афинитетот на HB до O 2.
6. Кооперативните промени во конформацијата на покровителите на хемоглобинот го забрзуваат врзувањето o 2 во белите дробови и враќањето на ткивото. Во белите дробови, високиот парцијален притисок O 2 придонесува за врзувањето на тоа со HB во активниот центар на четири протеар (2α и 2β). Активниот центар на секој протеер, како и во миоглобинот, се наоѓа помеѓу две α-спирали (F и E) во хидрофобниот џеб. Таа содржи не-pker дел - ГЕМ прикачен на протеинскиот дел со множество на слаби хидрофобни интеракции и една издржлива врска помеѓу FE 2 + GEMA и GIS F8 (види слика 1.21).
Во деоксихемоглобинот, благодарение на оваа врска со ГИС Ф 8, атомот Fe 2 + е од авионот на Хем кон хистидин. Обврзувањето O 2 со Fe 2 + се јавува на другата страна на Хемот во областа ГИС Е 7 користејќи единствена бесплатна координација. ГИС Е 7 обезбедува оптимални услови за врзување o 2 со Heme Irone.
Додавањето на O 2 до Fe +2 еден-заштитник Atom го предизвикува своето движење на авионот на Хем, а остатокот од хистинејата се поврзува со него
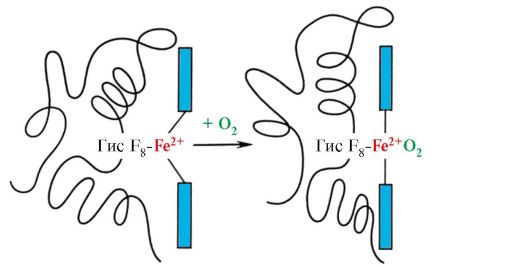 Сл. 1.23. Промени во конформацијата на околините хемоглобин кога се поврзани со O 2
Сл. 1.23. Промени во конформацијата на околините хемоглобин кога се поврзани со O 2
Ова доведува до промена во конформацијата на сите полипептински синџири поради нивната конформациона личност. Промената во конформацијата на другите синџири ја олеснува нивната интеракција со следните молекули околу 2.
Четвртата молекула О 2 се приклучува на хемоглобин 300 пати полесно од првата (слика 1.24).
 Сл. 1.24. Кооперативни промени во конформацијата на покровителите на хемоглобин во неговата интеракција со О 2
Сл. 1.24. Кооперативни промени во конформацијата на покровителите на хемоглобин во неговата интеракција со О 2
Во ткивата, секоја следна молекула е полесна од претходната, исто така поради кооперативни промени во конформацијата на демонстрантите.
7. CO 2 и H +, кои се формираат за време на катаболизмот на органските супстанции, го намалуваат афинитетот на хемоглобинот до О 2 пропорционално на нивната концентрација. Енергијата потребна за работењето на клетките се произведува главно во Mitochondria при оксидирање на органски супстанции кои користат O 2 испорачани од лесен хемоглобин. Како резултат на оксидацијата на органски супстанции, се формираат финални производи од нивното распаѓање: CO 2 и k 2 O, чиј број е пропорционален на интензитетот на оксидационите процеси.
CO 2 дифузијата излегува од клетките во крвта и продира во еритроцитите, каде што, под дејство на ензимот Carbanthidase се претвора во коалиска киселина. Оваа слаба киселина дисоцијација на протон и јонски бикарбонат.
N + способен да се приклучи на радикалите ГИС 14 6 во α- и β-синџири на хемоглобин, т.е. Во области далечински од полите. Прототацијата на хемоглобинот го намалува својот афинитет кон O 2, придонесува за расцепување на O 2 на Oxyne, формирање на деоксинев и го зголемува протокот на кислород во ткивото пропорционално на бројот на протони (Слика 1.25).
Зголемувањето на износот на ослободениот кислород, во зависност од зголемувањето на концентрацијата на H + во еритроцити, се нарекува ефект на Борон (наречен дански физиолог Кристијан Бор, кој прв го открил овој ефект).
Во белите дробови, високиот парцијален притисок на кислород придонесува за неговото врзување за Disoxyn, со што се намалува афинитетот на протеинот до H +. Опуштените протони под дејство на карбантидази комуницираат со бикарбонати со формирање на CO 2 и H2
 Сл. 1.25. Зависноста на афинитетот на HB до O 2 за концентрацијата на CO 2 и протоните (ефект на бора):
Сл. 1.25. Зависноста на афинитетот на HB до O 2 за концентрацијата на CO 2 и протоните (ефект на бора):
Но,- Влијание на концентрацијата на CO 2 и H + на пуштањето на 2 од комплексот со HB (ефект на BOHR); Б.- Оксигенација на деоксихемоглобин во белите дробови, образованието и распределбата на CO 2.
Како резултат на CO 2 влегува во алвеоларниот простор и се отстранува со издишаниот воздух. Така, износот на хемоглобин кислород ослободен во ткивата е регулиран со катаболни производи на органски супстанции: поинтензивно распаѓање на супстанции, на пример, за време на физичкиот напор, толку е поголема концентрацијата на CO 2 и H + и поголем кислород се добива од ткивото како резултат на намалувањето на афинитетот на HB до O 2.
8. Алостерична регулација на HB афинитет до 2 лиганд - 2,3-бисфосфоглицерски.Во црвените крвни клетки од производот за оксидација на гликоза - 1,3-бисфосфоглицерски се синтетизира со хемоглобин од алто-мобилен лига - 2,3-бисфосфоглицерат (2,3-bfg). Под нормални услови, концентрацијата на 2,3-BFG е висока и споредлива со концентрацијата на HV. 2,3-bfg има силен негативен полнеж -5.
 Бисфосфоглицерат во ткивните капилари, обврзувачки за деоксиемоглобин, го зголемува приносот на кислородот во ткивото, намалувајќи го афинитетот на HB до O 2.
Бисфосфоглицерат во ткивните капилари, обврзувачки за деоксиемоглобин, го зголемува приносот на кислородот во ткивото, намалувајќи го афинитетот на HB до O 2.
Во центарот на тетрамер хемоглобин молекула е празнина. Ги формира остатоците од аминокиселините на сите четири протераници (види слика 1.22). Во капиларите на ткивата, прототацијата на HB (ефектот на бор) води кон прекин на комуникацијата помеѓу HEME и 2 жлезда. Во молекула
дексихемоглобинот во споредба со оксимемоглобинот. Дополнителни јонски обврзници се јавуваат, кои ги поврзуваат демонстрантите, како резултат на што се зголемува големината на централната празнина во споредба со оксимимоглобинот. Централната празнина е 2,3-БФГ се приклучи на место за хемоглобин. Поради разликите во големината на централната празнина, 2,3-BFG може да се приклучи само од деоксихемоглобин.
2,3-bfg комуницира со хемоглобин во заговор отстранети од активните протеински центри и припаѓа алостеричен(регулаторни) лиганди, и централната празнина на HB е алостеричен центар.2.3-BFG има силен негативен полнеж и комуницира со пет позитивно наелектризирани групи од два β-синџири од HB: N-терминал α-амино група вратило и Лиз радикали 82 ГИС 143 (Слика 1.26).
 Сл. 1.26. БГГ во централната празнина на деоксихемоглобин
Сл. 1.26. БГГ во централната празнина на деоксихемоглобин
BFG се врзува за три позитивно наелектризирани групи во секој β-синџир.
Во капиларите на ткивата, деоксихемоглобинот во интеракција со 2,3-bfg и помеѓу позитивно наелектризирани β-ладински радикали и негативно наелектризирани лиганд формирани јонски обврзници кои ја менуваат сообразноста на протеинот и го намалуваат афинитетот на HB до O 2. Намалувањето на афинитетот на HB до O 2 придонесува за поефикасен излез од 2 во ткиво.
Во белите дробови на висок парцијален притисок, кислород комуницира со HB, приклучувајќи се на Glame Gland; Во овој случај, конформацијата на промените во протеините, централната празнина се намалува и 2,3-BFG од Алто-солидниот центар е раселено.
Така, олигомерните протеини имаат нови својства во споредба со мономерните протеини. Приложување на лиганди во области,
просторно отстранети едни од други (altowork), може да предизвика конформациски промени во целата протеинска молекула. Поради интеракцијата со регулаторните лиганди, конформацијата се менува и прилагодувањето на функцијата на протеинската молекула на еколошките промени.
Тема 1.5. Одржување на мајчин конформација на протеините во клеточните услови
Во клетките во процесот на синтеза на полипептидни синџири, нивните возила преку мембраните во соодветните клетки, за време на преклопувањето (формирање на мајчин конформација) и за време на собранието на олигомерни протеини, и во периодот на нивното функционирање Во структурата на протеините, посредни, склони кон агрегација, нестабилни конформации се јавуваат. Хидрофобни радикали, во мајчин конформација, обично скриени во молекулата на протеини, во нестабилната конформација се на површината и се стремат кон комбинирање со истите слабо растворливи групи во вода со групи други протеини. Во клетките на сите познати организми беа пронајдени специјални протеини, кои обезбедуваат оптимални фабрички клеточни протеини, стабилизирање на нивната мајчин конформација за време на работењето и, што е особено важно, ја поддржува структурата и функциите на интрацелуларните протеини со повреда на хоместазата. Овие протеини добија име "Хаперинс",што преведено од француски значи "Дадилка".
1. Молекуларни хапери и нивната улога во спречувањето на денатурацијата на протеините.
Shaperons (W) се класифицирани по тежина на субедити. Високата молекуларна тежина се шегуваше со маса од 60 до 110 kd. Меѓу нив се најубавите три класи: W-60, WC-70 и W-90. Секоја класа вклучува семејство на поврзани протеини. Така, составот на W-70 вклучува протеини со молекуларна тежина од 66 до 78 kd. Ниските молекуларни тежински шепери имаат молекуларна тежина од 40 до 15 kd.
Меѓу хапериите се разликуваат конститутивенпротеини, висока базална синтеза, чија не зависи од стресни ефекти врз клетките на телото и индуцибиленсинтезата за која под нормални услови е слаба, но остро се зголемува за време на стресни ефекти. Индуктивни шепери се нарекуваат "протеини на топлински шок", бидејќи тие првпат беа откриени во клетките кои се подложени на високи температури. Во клетките поради високата концентрација на протеини, спонтаната ренатација на делумно денатурираните протеини е попречена. WC-70 може да го спречи процесот на денатурација на денатурата и да го промовира реставрацијата на мајчин конформација на протеините. Молекуларни хаперизони-70- Класа на протеини со висока коло лоцирана во сите клетки на ќелијата: цитоплазма, јадро, ендоплазмичен ретикулум, митохондрии. На карбоксилниот крај на единствениот полипептид коло на W-70, постои заговор кој е жлеб способен за интеракција со пептиди
од 7 до 9 аминокиселински остатоци збогатени со хидрофобни радикали. Таквите области во глобуларни протеини се случуваат околу секои 16 амино киселини. WC-70 е способен за заштита на протеините од инактивацијата на температурата и да ја врати усогласеноста и активноста на делумно денатурираните протеини.
2. Улогата на ѕидовите во преклопните протеини.Во синтезата на протеините на рибозомот, регионот N-терминал на полипептидот се синтетизира пред Ц-терминалот. За да формираат мајчин конформација, неопходна е целосна аминокиселинска секвенца на протеини. Во процесот на синтеза на протеините на Sperlons-70, поради структурата на нивниот активен центар, областите на полипептидот склони кон агрегација, збогатени со хидрофобни радикали на аминокиселини пред завршувањето на синтезата (слика 1.27, а).
 Сл. 1.27. Учество на хаперизони во преклопни протеини
Сл. 1.27. Учество на хаперизони во преклопни протеини
А - учество на ShaperOne-70 во спречување на хидрофобни интеракции помеѓу делниците на синтетизираниот полипептид; Б - Формирање на мајчин протеински конформации во комплекс од чеперион
Многу високи молекуларни протеини кои имаат сложена конформација, како што е структурата на доменот, произведуваат преклопување во посебен простор формирана од W-60. Sh-60.функција во форма на олигомерен комплекс кој се состои од 14 подединици. Тие формираат два шупливи прстени, од кои секоја се состои од седум подединици, овие прстени се поврзани едни со други. Секоја подединица W-60 се состои од три домени: апикални (горе), збогатени со хидрофобни радикали со кои се соочуваат прстените, средната и екваторијалната празнина (слика 1.28).
 Сл. 1.28. Структурата на комплексот Shaperonin се состои од 14 W-60
Сл. 1.28. Структурата на комплексот Shaperonin се состои од 14 W-60
А-страничен поглед; Б - врвот поглед
Синтетизирани протеини кои имаат елементи на површината карактеристика на развиените молекули, особено хидрофобни радикали, спаѓаат во шуплината на прстените што се пренасочуваат. Во одредена средина на овие шуплини, постои биста на можни конформации, до единствената, енергијата е најкорисна (слика 1.27, б). Формирањето на конформации и ослободување на протеини е придружено со хидролиза на АТП во екваторијалниот регион. Обично таквото преклопување зависно од преклопување бара трошоците за значителен износ на енергија.
Покрај учеството во формирањето на тродимензионална структура на протеини и ренатоцијата на делумно денатурираните протеини, се неопходни за протокот на таквите фундаментални процеси како собранието на олигомерни протеини, препознавање и транспорт во лизозомите на денатурирани Протеини, протеински транспорт преку мембрани, учество во регулирањето на протеинските комплекси.
Тема 1.6. Различни протеини. Семејство на протеини на пример на имуноглобулини
1. Протеините играат клучна улога во виталната активност на поединечните клетки и целиот мултичелуларен организам, а нивните функции се изненадувачки различни. Ова се определува со особеностите на примарната структура и конформациите на протеините, уникатноста на структурата на активниот центар и способноста за поврзување на специфични лиганди.
Само многу мал дел од сите можни варијанти на пептидните синџири може да се земе стабилна просторна структура; најмногу
од нив, тоа може да потрае многу конформации со истата Gibbs енергија, но со различни својства. Примарната структура на мнозинството познати протеини избрани со биолошка еволуција обезбедува исклучителна стабилност на една од конформациите, со што се утврдуваат карактеристиките на функционирањето на овој протеин.
2. Семејни протеини.Во рамките на еден биолошки видови Замена на остатоци од амино киселини може да доведе до различни протеини кои вршат сродни функции и со хомологни секвенци на амино киселини. Таквите поврзани протеини имаат неверојатно слична конформација: бројот и интерпозицијата на α-хеликс и (или) β-структури, поголемиот дел од вртежите и свиоците на полипептидни синџири се слични или идентични. Протеините со хомологни делови од полипептидниот синџир, слична конформација и сродни функции се изолирани во семејството на протеини. Примери на семејства на протеини: серински протеини, семејство на имуноглобулини, семејството Миглобин.
Серински протеиназа- семејство на протеини кои ја извршуваат функцијата на протеолитички ензими. Тие вклучуваат дигестивни ензими - chymotrypsin, трипсин, еластаза и многу коагулациски фактори. Овие протеини имаат 40% од одредбите на идентични амино киселини и многу блиска конформација (слика 1.29).

Сл. 1.29. Просторни структури на еластаза (а) и chymotrypsin (б)
Некои супстанции за амино киселини доведоа до промена на специфичноста на подлогата на овие протеини и појавата на функционален колектор во семејството.
3. Семејство на имуноглобулини.Во работата на имунолошкиот систем, протеините на суперфамилија имуноглобулинов играат огромна улога, која вклучува три семејства на протеини:
Антитела (имуноглобулини);
T-Lymphocyte рецептори;
Протеините на главната хистокомпатибилност комплекс - главен хистокомпен комплекс).
Сите овие протеини имаат структура на доменот, се состојат од хомологни домени слични на имунопород и вршат слични функции: комуницираат со туѓи структури или растворени во крв, лимфа или меѓуцентрална течност (антитела), или на површината на клетките (сопствени или странци) .
4. Антитела- специфични протеини произведени од лимфоцити како одговор на влегувањето во телото на странска структура наречена антиген.
Карактеристиките на структурата на антителата
Наједноставните молекули на антитела се состојат од четири полипептински синџири: две идентични бели дробови - L со околу 220 амино киселини и две идентични тешки H, кои се состојат од 440-700 амино киселини. Сите четири синџири во молекулата на антитела се поврзани со плуралноста на не-ковалентни обврзници и четири дисулфидни обврзници (слика 1.30).
Светлосните синџири на антителото се состојат од два домени: променлива (VL), која се наоѓа во N-терминалниот регион на полипептидниот синџир и константа (CL), кој се наоѓа на крајот. Тешките синџири обично имаат четири домени: една променлива (VH), која се наоѓа на N-крајот, и три постојани (CH1, CH2, SNZ) (види слика 1.30). Секој домен на имуноглобулин има β-преклопена надградба, во која два цистеински остатоци е поврзан со дисулфидна врска.
Помеѓу двата постојани домени на CH1 и CH2 постои парцела која содржи голем број на пролин остатоци кои го попречуваат формирањето на секундарната структура и интеракцијата на соседните Х-синџири на овој сегмент. Оваа област на шарка ја дава молекулата на флексибилноста на антителата. Постојат две идентични антиген-обврзувачки парцели помеѓу променливите домени на тешки и лесни синџири (активни центри за врзување на антиген), така што таквите антитела често се нарекуваат бивалент.Во врзувањето на антигенот со антитело, не е вклучена целата аминокиселинска секвенца на варијабилни делови од двата синџири на двата синџири, а само 20-30 амино киселини се наоѓаат во хиперваливите региони на секој синџир. Токму овие области ја одредуваат уникатната способност на секој тип на антитело за да комуницираат со соодветниот комплементарен антиген.
Антителата се една од линиите за заштита од телото против имагинарните туѓи организми. Нивното функционирање може да се подели во две фази: првата фаза е препознавањето и врзувањето на антигенот на површината на туѓите организми, што е можно поради присуството на антитела врз основа на антиген-обврзувачки во структурата; Втората фаза е иницирање на процесот на неактивност и уништување на антигенот. Специфичноста на втората фаза зависи од класата на антитела. Постојат пет класи на тешки синџири кои се разликуваат едни од други на структурата на постојаните домени: α, δ, ε, γ и μ, во согласност со која се разликуваат петте класи на имуноглобулини: A, D, E, G и M .
Карактеристиките на структурата на тешките синџири се прикачени на шаркираните области и C-терминалните области на тешки синџири карактеристични за секоја класа конформација. По обврзувањето на антигенот со антитело, конформационите промени во постојаните домени ја дефинираат патеката за отстранување на антигенот.
 Сл. 1. 30. ИГГ Структура на доменот
Сл. 1. 30. ИГГ Структура на доменот
Имуноглобулини М.
Имуноглобулините М имаат две форми.
Мономерна форма- 1. Класа на антитела произведени со развој во лимфоцити. Подоцна, многу Б-клетки се префрлија на производство на други класи на антитела, но со истата активност за врзување на антиген. ИГМ е вграден во мембраната и ја врши улогата на антигенски рецептор. Вградување IGM во клеточната мембрана е можно поради присуството на хидрофобни аминокиселински остатоци во опашката.
Секретор на ИГМ.содржи пет мономер подедини поврзани со едни со други дисулфид обврзници и дополнителен полипептид J-синџир (Слика 1.31). Тешките мономерни синџири на оваа форма не содржат хидрофобна опашка. Пенталот има 10 обврзувачки центри со антиген и затоа е ефективен во признавањето и отстранувањето на првиот антиген во телото. Секреталната форма ИГМ е главната класа на антитела кои се секретираат во крвта за време на примарен имунолошки одговор. ИГМ Врзувањето со антиген ја менува сообразноста на ИГМ и го поттикнуваше врзувањето на првата протеинска компонента на системот за дополнување (системот за комплементот е збир на протеини кои се вклучени во уништувањето на антигенот) и активирањето на овој систем. Ако антигенот се наоѓа на површината на микроорганизмот, системот за дополнување предизвикува повреда на интегритетот на клеточната мембрана и смртта на бактериска клетка.
Имуноглобулини G.
Во квантитативно, оваа класа на имуноглобулини преовладува во крвта (75% од сите ИГ). Igg - мономери, главната класа на антитела излачувани во крвта за време на секундарен имунолошки одговор. По интеракцијата на IgG со површински антигени микроорганизми, комплексот за антиген-антитела е способен да ги поврзе и активира протеините на системот за комплементот или може да комуницира со специфични макрофаги и неутрофилни рецептори. Интеракција со фагоцити води
 Сл. 1.31. Структура на секреторната форма ИГМ
Сл. 1.31. Структура на секреторната форма ИГМ
на апсорпција на комплексите за антиген-антитела и нивно уништување во клетките на клетките. IgG е единствената класа на антитела кои можат да навлезат во плаценталната бариера и да обезбедат интраутерина заштита на фетусот од инфекции.
Имуноглобулини А.
Главната класа на антитела присутни во тајни (млеко, плунка, тајните на респираторниот тракт и цревниот тракт). ИГА тајни главно во димерна форма, каде што мономерите се поврзани едни со други преку дополнителен J-синџир (слика 1.32).
ИГА не комуницира со системот за дополнување и фаицитифични клетки, но, обврзувачки за микроорганизми, антитела ги спречуваат пристапувањето кон епителните клетки и пенетрација во телото.
Имуноглобулини Е.
Имуноглобулините Е се претставени од мономери во кои тешките ε-синџири содржат, како и μ-синџири на имуноглобулин М, една променлива и четири постојани домени. IGE по секрецијата се врзуваат за нив
 Сл. 1.32. Структурата на IGA.
Сл. 1.32. Структурата на IGA.
C-терминални делови со соодветни рецептори на површината на дебели клетки и базофили. Како резултат на тоа, тие стануваат рецептори за антигени на површината на овие клетки (слика 1.33).
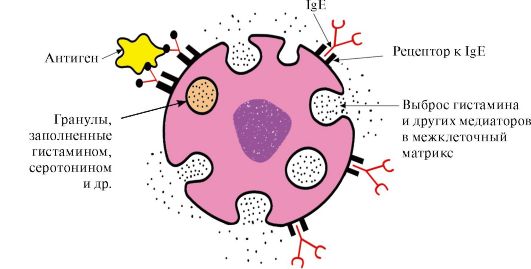 Сл. 1.33. IGE интеракција со антигенот на површината на масните клетки
Сл. 1.33. IGE интеракција со антигенот на површината на масните клетки
По прицврстувањето на антигенот е прикачен на соодветните антиген-обврзувачки делови на ИГЕ, клетките добиваат сигнал до секреција на биолошки активни супстанции (хистамин, серотонин), кои во голема мера се одговорни за развој на воспалителен одговор и за Манифестација на такви алергиски реакцииКако астма, уртикарија, поленска треска.
Имуноглобулини D.
Имуноглобулините D се откриени во серумот во многу мала количина, тие се мономери. Во тешки δ-синџири постојат една променлива и три постојани домени. Игда ја врши улогата на при-лимфоцити, други функции се уште се непознати. Интеракцијата на специфични антигени со рецептори на површината на Б-лимфоцитите (IGD) доведува до пренос на овие сигнали во ќелијата и вклучување на механизми кои обезбедуваат репродукција на овој клон на лимфоцити.
Тема 1.7. Физичко-хемиски својства на протеини и методи на нивно одвојување
1. Индивидуалните протеини се разликуваат во физичко-хемиските својства:
Молекули;
Молекуларна тежина;
Вкупно полнење, чија вредност зависи од односот на анјонски и катјонски групи на амино киселини;
Односот на поларните и не-поларни радикали на амино киселини на површината на молекулите;
Степенот на отпорност на ефектите на различни агенти за денатурирање.
2. Зависини од растворливост зависисвојствата на протеините наведени погоре, како и на составот на медиумот во кој се раствора протеинот (pH вредности, составот на сол, температура, присуство на други органски супстанции способни за интеракција со протеинот). Големината на обвинението за молекули на протеини е еден од факторите кои влијаат на нивната растворливост. Ако загубата на полнење во изоелектричната точка, протеините полесно се полесни и паѓаат во талог. Ова е особено карактеристично за денатурираните протеини, во кои радикалите на хидрофобните амино киселини се на површината.
На површината на молекулата на протеини, постојат и позитивни и негативно наелектризирани амино киселински радикали. Бројот на овие групи, и, следствено, вкупниот полнеж на протеините зависи од pH на медиумот, т.е. Односот на концентрацијата на H + - и тоа е -groups. Во кисела срединазголемувањето на концентрацијата на H + доведува до сузбивање на дисоцијација на карбоксилни групи - + - + H +\u003e - Кокси и намалување на негативниот полнеж на протеини. Во алкален медиум, врзувањето на вишокот од ИТ-протони формирани за време на дисоцијацијата на амино групи -NH3 + + IT - - NH 2 + H 2 O со формирање на вода води кон намалување на позитивниот полнеж на протеините . PH вредноста во која протеинот има вкупна нула задолжен, наречен изоелектрична точка (ИЕТ).Во ИЕТ, бројот на позитивни и негативно наелектризирани групи е подеднакво, односно Протеинот е во изоелектрична состојба.
3. Поделба на поединечни протеини.Карактеристиките на структурата и функционирањето на телото зависат од збир на протеини кои се синтетизираат во неа. Студијата за структурата и својствата на протеините е невозможно без нивно ослободување од ќелијата и прочистувањето од други протеини и органски молекули. Фази на селекција и прочистување на поединечни протеини:
Уништување на клеткитеткивото се изучува и добива хомогени.
Одвојување на хомогенирање на фракциицентрифугирање, добивање на нуклеарна, митохондријална, цитозолна или друга фракција која го содржи саканиот протеин.
Изборната термална денатурација- Краткорочно загревање на протеинското раствор, на кое можете да го отстраните дел од нечистотиите на протеини (ако протеинот во однос на термостата).
Садење.Различни протеини спаѓаат во талог на различни концентрации на сол во растворот. Постепено, зголемување на концентрацијата на сол, може да се добие голем број на поединечни фракции со доминантна содржина на протеинот објавен во еден од нив. Најчесто се користи фракционирање на протеини, се користи амониум сулфат. Протеините со најмалата растворливост паѓаат во талог на мали концентрации на солени.
Гел филтрација- Метод на миеринг молекули преку отечени гранули на сефадекс (три-димензионални полисахаридцишици, кои имаат пори). Брзината на поминување на протеините преку колона исполнета со сефадекс ќе зависи од нивната молекуларна тежина: толку помалку тежина на протеинските молекули, толку полесно ги навлезат во внатрешноста на гранулите и подолго одложено, толку е поголема масата од колоната.
Ultracentrifugation.- Методот што се состои во фактот дека протеините во цевката центри се ставаат во роторот Ultracentifuge. Кога ротирачкиот ротор, стапката на седиментација на протеините е пропорционална на нивната молекуларна тежина: фракциите на потешки протеини се поблиску до дното на цевката за тестирање, полесни - поблиску до површината.
Електрофореза- методот врз основа на разликите во брзината на движење на протеините во електричното поле. Оваа големина е пропорционална на полнењето на протеините. Електрофореза протеините се изведуваат на хартија (во овој случај, брзината на движење на протеините е пропорционална само на нивното полнење) или во полиакриламид гел со одредена количина на пори (брзината на движење на протеините е пропорционална на нивното полнење и молекуларна Тежина).
Јонска размена на хроматографија- метод на фракционирање врз основа на обврзувачки јонизирани групи на протеини со спротивставени групи на минијаци на јонска размена (нерастворливи полимерни материјали). Протеините врз основа на протеини со смола е пропорционална на пакетот на протеини. Протеините adsorbed на јонски размена на полимер може да се мијат со nacl решенија со зголемени концентрации; Помалку протеински полнеж, помалата концентрација на Nacl ќе биде потребна за измивање на протеинот поврзан со јонските групи на смолата.
Афинитет хроматографија- Најфицифичен метод на изолирање на индивидуалните протеини. Инертниот полимер е ковалентно придружуван од лиганд на било кој протеин. Кога решението на тимот е пренесен низ колоната со полимер поради комплементарно врзување на протеин на протеин со лиганд на колоната, само специфичен протеин-специфичен протеин е адсорбиран.
Дијализа- Методот што се користи за отстранување на соединенија со ниска молекуларна тежина од раствор на избраниот протеин. Методот се заснова на неможноста на протеините да помине низ полупропустлива мембрана, за разлика од ниските молекуларни тежини. Се користи за прочистување на протеините од ниско молекуларна тежина нечистотии, како што се соли по садењето.
Задачи за воннаставна работа
1. Пополнете ја табелата. 1.4.
Табела 1.4. Компаративна анализа на структурата и функциите на поврзаните протеини - миглобин и хемоглобин
а) Потсетиме на структурата на активниот центар на МВ и Х.Б.. Каква улога играат хидрофобните амино киселински радикали во формирањето на активни центри на овие протеини? Опишете ја структурата на активниот центар на МВ и Хи и механизмите на приврзаност кон него лиганди. Каква улога е остатоците од ГИС Ф 8 и ГИС Е 7 во функционирањето на активниот центар на МВ.
б) Кои нови својства се тесно поврзани олигомерни протеини во споредба со мономерниот миоглобин - хемоглобин? Објаснете ја улогата на кооперативните промени во конформацијата на демонстрантите во молекулата на хемоглобинот, ефектот на концентрациите на CO 2 и протоните во афинитетот на хемоглобинот до кислород, како и улогата на 2,3-BFG во алт-користењето функцијата на функцијата NC.
2. Дајте карактеристика на молекуларните садови, обрнувајќи внимание на поврзувањето на нивната структура со функцијата.
3. Кои протеини се обединети во семејствата? Користејќи го примерот на семејството на имуноглобулини, ги утврдува сличностите на структурата и поврзаните функции на протеините на ова семејство.
4. Често, се потребни прочистени индивидуални протеини за биохемиски и медицински цели. Објаснете дека физичко-хемиските својства на протеините се засноваат на методите на нивното одвојување и чистење.
Задачи за самоконтрола
1. Изберете ги вистинските одговори.
Функции на гемоглобинот:
А. Транспорт на 2 од белите дробови во ткиво Б. Транспорт N + од ткаенини во белите дробови
Б. Одржување на постојаноста на крв PH на градот Транспорт CO 2 од белите дробови во ткаенината
Д. Транспорт CO 2 од ткаенините во белите дробови
2. Изберете ги вистинските одговори. Лигандα -Протомер NV е:А. ГЕМ.
Б. Кислород
Б. 2,3-БФГ
Д. β-правилно
3. Изберете ги вистинските одговори.
Хемоглобинот во контраст со миглобинот:
A. Има кватернерна структура
Б. секундарна структура е претставена со α-спирали
Б. се однесува на сложени протеини.
Г. Интеракција со алостеричен лиганд Д. ковалентно поврзан со полите
4. Изберете ги вистинските одговори.
Афинитетот на HB до 2 се намалува:
A. Кога прицврстувате една молекула околу 2 Б. Кога се расклопувате една молекула o 2
Б. Кога комуницирате со 2,3-bgg
Г. Кога е поврзан со протометарот H + D. со намалување на концентрацијата од 2,3-bfg
5. Постави натпревар.
За видовите на HB, тоа е карактеристично:
A. Deoxy Form Forms Фибриларни единици Б. Содржи во составот на два α- и два δ-синџири
Б. Преовладувачката форма на HB во еритроцитите на возрасен човек во активниот центар содржи дијамант со Fe + 3
Д. содржи две α- и два γ-синџири 1. NVA 2.
6. Постави натпревар.
Ligands NV:
А. се врзува за HB во Al Motor Center
Б. има многу висок афинитет за активниот центар на HB
Б. Со поврзување, го зголемува афинитетот на HB до O 2 оксидира Fe + 2 во Fe + 3
Д. формира ковалентна врска со GISH8
7. Изберете ги вистинските одговори.
Chaperons:
А. Протеини присутни во сите клетки на ќелијата
Б. Синтезата е подобрена во стресни ефекти.
Б. Учествувајте во хидролиза на денатурираните протеини
Г. се вклучени во одржувањето на конформацијата на мајчин протеини
Д. Креирај органелс во кои се формира конформација на протеините
8. Инсталирајте го натпреварот. Имуноглобулини:
А. Секреталната форма има форма на Пентар
Б. Класа И пенетрантска бариера
Б. ИГ - рецептор на масни клетка
Г. Главна класа ИГ присутна во тајните на епителните клетки. D. B-Lymphocyte рецептор, чија активација обезбедува репродукција на клетки
9. Изберете ги вистинските одговори.
Имуноглобулини Е:
А. Макрофаги Б. се произведуваат со тешки ε-синџири.
Б. Вграден во мембраната T-Lymphyte
G. Направете улога на мембранските антигени рецептори на масни клетки и базофила
Г. Одговорен за манифестација на алергиски реакции
10. Изберете ги вистинските одговори.
Методот на поделба на протеините се базира на разлики во нивната молекуларна тежина:
А. Гел филтрација
Б. Ultracentrifugation.
Б. Електрофореза кај полиакриламид гел јон размена на хроматографија
Д. афинитет хроматографија
11. Одбери го точниот одговор.
Методот на поделба на протеините се базира на разлики во нивната растворливост во водата:
А. Гел филтрација Б. Садење
Б. јон размена хроматографија на афината хроматографија
Д. Електрофореза во полиакриламид гел
Стандарди за одговори на "работни места за самоконтрола"
1. А, Б, Б, Д
2. А, Б, Б, Д
5. 1-б, 2-А, 3-г
6. 1-б, 2-б, 3-а
7. А, Б, Г, Д
8. 1-g; 2-б, 3-во
Главни термини и концепти
1. Олигомерни верверички, соодветна, кватернерна протеинска структура
2. Кооперативни промени во конформацијата на демонстрантите
3. БОН ЕФЕКЦИЈА
4. Алостерична регулација на функциите на протеините, алостеричниот центар и алостеричен ефектор
5. Молекуларни хапери, топлински шок протеини
6. Семејни протеини (серински протеани, имуноглобулини)
7. IGM-, G-, Е-, А-обврзница со функција
8. Резиме на протеини, протеински изоелектрична точка
9. Електрофореза
10. Пеење
11. Гел филтрација
12. јонска размена на хроматографија
13. Ultracentrifugation
14. Афинарна хроматографија
15. Електрофореза на крвни плазма протеини
Задачи за ревизорска работа
1. Споредете ги зависностите од степени на сатурација на хемоглобинот (HB) и Myoglobin (MB) кислород од нејзиниот парцијален притисок во ткивата
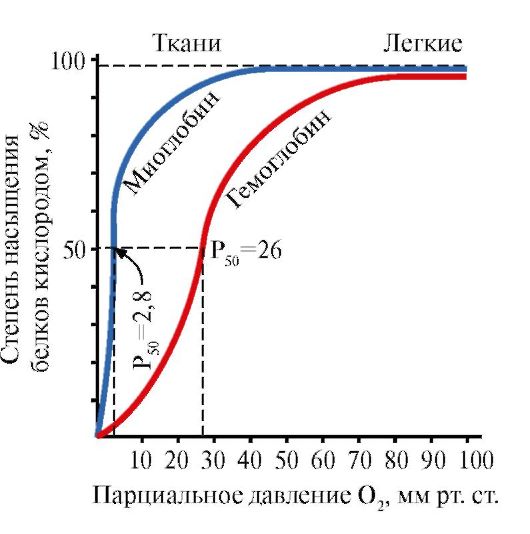 Сл. 1.34. Зависноста на сатурацијата на МВ иЊујорккислород од својот парцијален притисок
Сл. 1.34. Зависноста на сатурацијата на МВ иЊујорккислород од својот парцијален притисок
Ве молиме имајте предвид дека обликот на крива на кислород од протеини е различен: за миоглобинот - хипербола, за хемоглобин - сигмоидна форма.
1. Споредете ги вредностите на делумниот кислороден притисок, во кој MB и HV се заситени со 2 за 50%. Кој од овие протеини се карактеризира со повисок афинитет за 2?
2. Кои карактеристики на МВ структурата го одредуваат својот висок афинитет за 2?
3. Какви карактеристики на структурата на НВ му овозможуваат да дадат 2 во капиларите на ткивата за одмор (со релативно висок парцијален притисок од 2) и драматично го зголемуваат ова враќање во работните мускули? Кој својство на олигомерни протеини го обезбедува овој ефект?
4. Пресметајте колку количината од 2 (во%) дава оксигенирана хемоглобин одмор и работа на мускулите?
5. Направете заклучоци за поврзувањето на протеинската структура со својата функција.
2. Износот на кислород ослободен од хемоглобинот кај капиларите зависи од интензитетот на процесите на катаболизмот во ткивата (ефект на бор). Како промените во метаболизмот во ткивата го регулираат афинитетот на Њујорк до О 2? Ефект на CO 2 и H + на афинитет на Њујорк до 2
 1. Опишете го ефектот на бор.
1. Опишете го ефектот на бор.
2. Во која насока процесот претставен во шемата продолжува:
а) во капиларите на белите дробови;
б) Капилари за искачување?
3. Во Че. физиолошко значење Ефект на БОХР?
4. Зошто интеракцијата на NV со H + во области оддалечени од полите, го менува афинитетот на протеинот до 2?
3. Афинитетот на НБЕ до О2 зависи од концентрацијата на нејзиниот лиганд - 2,3-бифосфо-глицерат, која е регулатор на Alto пушењето на афинитетот на HV до O 2. Зошто интеракцијата на лиганд во далечинскиот управувач од активниот центар влијае на функцијата на протеини? Како 2,3-bfg го регулира афинитетот на Њујорк до 2? За да ја решите задачата, одговорете на следниве прашања:
1. Каде и од кои 2,3-бифосфоглицери (2,3-bfg) се синтетизира? Напишете го на формулата, наведете го полнењето на оваа молекула.
2. Со која форма на хемоглобин (окси или деок) го поврзува БФГ и зошто? Во кој дел од молекулата на ХБ е интеракција?
3. Во која насока процесот презентиран во шемата продолжува
а) во капилари на ткаенина;
б) во капиларите на белите дробови?
4. Каде треба да биде повисока концентрација на комплексот
HB-2,3-BFG:
а) во капиларите на мускулите во состојбата на одмор
б) во капиларите на работните мускули (обезбедена иста концентрација на БФГ во црвените крвни клетки)?
5. Како ќе се смени афинитетот на HV во кислород кога ќе се прилагоди на лицето на условите на висорамнините, ако концентрацијата на BFG во еритроцитите се зголемува? Кое е физиолошкото значење на овој феномен?
4. Уништувањето на 2.3-БГГ за време на складирањето на конзервирана крв ги нарушува функциите на НВ. Како афинитетот на HV се менува во O 2 во конзервирана крв, ако концентрацијата од 2,3-Bfg во црвени крвни клетки може да се намали од 8 до 0,5 mmol / l. Дали е можно да се прелева како крв на сериозно болни пациенти ако концентрацијата од 2,3-bfg е вратена не порано отколку за три дена? Дали е можно да се додаде 2,3-bfg на крвта за да се вратат функциите на еритроцитите?
5. Потсетиме на структурата на наједноставните молекули на имуноглобулин. Каква улога играат имуноглобулините во работата на имунолошкиот систем? Зошто ИГ често се нарекува бивалентен? Како е IG структура поврзана со нивната функција? (Опишете пример за било која класа на имуноглобулини.)
Физичко-хемиски својства на протеини и методи за нивно одвојување.
6. Како сенката на протеините влијае на растворливоста?
а) го одредува вкупниот пептид на PH 7
Ala-glu-tre-asp-liz cis
б) Како да го промените цената на овој пептид на pH\u003e 7, pH<7, рН <<7?
в) Која е изоелектричната точка на протеинот (ИЕТ) и во која средина лежи
На овој пептид?
г) Со тоа што значењето на pH вредноста ќе биде забележано најмалата растворливост на овој пептид.
7. Зошто кисело млеко за разлика од свежо во вриењето "преклопено" (т.е. млеко протеини казеин паѓа во седиментот)? Во свежото млеко, молекулите на казеин имаат негативно полнење.
8. За одвојување на поединечни протеини, се користи методот на галтрација. Мешавина што содржи протеини А, Б, Ц со молекуларни маси еднакви на 160.000, 80.000 и 60.000, соодветно, беше анализирана со гел филтрација (Слика 1.35). Пелетите на отечениот гел пропустливи за протеини со молекуларна тежина од помалку од 70.000. Кој принцип се заснова на овој метод за одвојување? Која од графиконите правилно ги одразува резултатите од фракционата? Наведете го редот на излезот на протеините A, B и C од колоната.
 Сл. 1.35. Користење на методот на филтрирање на гел за одвојување на протеини
Сл. 1.35. Користење на методот на филтрирање на гел за одвојување на протеини
9. На Сл. 1.36, и го покажува дијаграмот на електрофореза на хартиени протеини протеини на здрава личност. Релативните количини на протеински фракции добиени со користење на овој метод се: Албумин 54-58%, α 1 -глобулини 6-7%, α 2 -глебулини 8-9%, β-глобулини 13%, γ-глобулини 11-12%.
 Сл. 1.36 Електрофореза на хартиени плазма протеини на здрава личност (и) и пациент (б)
Сл. 1.36 Електрофореза на хартиени плазма протеини на здрава личност (и) и пациент (б)
I-γ-глобулини; II - β-глобулини; III -α 2-глобулин; IV -α 2-глобулин; V - Албумин
Многу болести се придружени со квантитативни промени во составот на серумските протеини (дисломисминија). Природата на овие промени се зема предвид при дијагностицирање и проценка на сериозноста и фазата на болеста.
Со помош на податоците дадени во табелата. 1.5, направи претпоставка за болеста за која електрофоретичниот профил презентиран на Сл. 1.36.
Табела 1.5. Промена на концентрацијата на серумските протеини во патологијата
Активен центар верверица -ова е центар за врзување на протеини со лиганд. Заговор е формиран на површината на глобусот, кој може да приложи други молекули наречени лиганди . Активниот центар на протеинот е формиран од страничните групи на амино киселини, најблиску на ниво на терциерна структура. Во линеарната секвенца на пептидниот синџир, тие можат да бидат на далечина далечински оддалеченост едни од други. Протеините покажуваат висока специфичност при интеракција со лиганд. Високата специфичност на интеракцијата на протеините со лигандот е обезбедена со комплементарност на структурата на активниот центар на протеинската структура на лиганд. Комплементација - Тоа е просторно и хемиско совпаѓање на интерактивните молекули. Центрите за врзување на протеини со лиганд често се наоѓаат помеѓу домените (на пример, Центарот за врзување на Tripsin со својот лиганд има 2 домени одделени со фурода).
Функционирањето на протеините се базира на нивната специфична интеракција со лиганди. 50.000 индивидуални протеини кои содржат уникатни активни центри способни да контактираат само специфични лиганди и, благодарение на карактеристиките на активната централна структура, за да ги прикажат карактеристичните функции. Очигледно, примарната структура содржи информации за функцијата на протеини.
Кватернерна структура - Ова е највисоко ниво на структурна организација, не е можно во сите протеини. Под кватернерната структура, разбраниот метод на поставување во просторот на полипептидни синџири и формирање на единствен во структурните и функционалните односи на макромолекуларно образование е разбрана. Секој одделно се зема полипептид синџир наречен прототерала или судиет , најчесто нема биолошка активност. Оваа способност на протеинот стекнува со одреден метод на просторно здружение на демонстранти вклучени во неговиот состав. Како резултат на молекулата се нарекува повикана олигомер (мултимер) .
Кватерната структура ќе ги стабилизира не-вирулентните врски кои се јавуваат помеѓу локациите за контакт на демонстрантите, кои комуницираат едни со други со видот на комплементарност.
Протеините кои имаат кватернерна структура вклучуваат многу ензими (лактат дехидрогеназа, глутамидхидрогеназа, итн.), Како и хемоглобин, контралективен мускулен протеин моцин. Некои протеини имаат мал број на сублинци 2 - 8, други стотици, па дури и илјадници подединици. На пример, тутунскиот мозаик вирус протеин има 2130 подединици.
Типичен пример за протеин кој има кватернерна структура е хемоглобин. Молекулата на хемоглобинот се состои од 4 подедини, трајни синџири на полипептид, од кои секоја е поврзана со дијаматот, од кои 2 полипептидни синџири се нарекуваат -2Afel и -2бет, тие се разликуваат во примарната структура и должината на полипептидниот синџир.
Комуникации формирање на кватернерна структура помалку издржлива. Под влијание на одредени агенти, протеинот е одделен на посебни подединици. Кога се отстранува агенсот на субедит, биолошката функција на протеинот повторно се обновува. Значи, кога хемоглобинот на уреата се додава во решение, се распаѓа во 4 компоненти на своите субедизи, кога уреата ја отстранува структурната и функционална улога на хемоглобинот е обновена.
Крај на работа -
Оваа тема спаѓа во делот:
Биохемија. Протеини. Амино киселини - структурни компоненти на протеини
Аминокиселински протеини Структурни компоненти на протеини .. Протеини. Протеините се азот со висока молекуларна тежина органски соединенија кои се состојат од амино киселини поврзани во синџирот.
Ако ви треба дополнителен материјал на оваа тема, или не сте го пронајдоле она што го барале, препорачуваме користење на пребарување за нашата работна база:
Она што ќе го направиме со добиениот материјал:
Ако овој материјал се покажа како корисен за вас, можете да го зачувате на вашата страница за социјално вмрежување:
| Tweet. |
Сите теми од овој дел:
Механизмот на дејство на ензими
Според современите идеи, кога ензимот комуницира со подлогата, можно е да се разликуваат 3 етапи: 1 фаза се карактеризира со дифузија на подлогата до Феррен
Киселина-главна катализа
Во активниот центар на ензимот содржи групи киселини и примарен тип. Групите на киселина се расцепуваат H + и имаат негативен полнеж. Основни групи за тип се приклучуваат на H + и имаат поло
Но). Хипотеза на Фишер
Според него, постои строга почва совпаѓање на подлогата и активниот центар на ензимот. Според Фишер, ензимот е хард структура, а подлогата е како актер на активниот процент
Размена на јаглени хидрати
Размена на јаглени хидрати 1. Главните јаглени хидрати на животинскиот организам, нивната биолошка улога. 2. Трансформација на јаглени хидрати во органите на дигестивниот систем. 3. биосинтеза и распаѓање
Биолошка улога на јаглени хидрати
Биолошка улога на јаглени хидрати: 1. Енергија. Кога се разликува оксидација1 g од јаглени хидрати на конечни производи (CO2 и H2O), се разликува 4.1-KCAL на енергија. Јагленохидратите сочинуваат околу 60-70
Трансформација на јаглени хидрати во дигестивниот тракт
Конверзијата на јаглени хидрати во дигестивниот тракт со главната јагленохидратна храна за човечкото тело е: скроб, гликоген, сахароза, лактоза. Несреќа
Биосинтеза и дезинтеграција на гликоген
Биосинтеза и гликоген распаѓање во ткивата. Гликоген болести. Беше откриено дека гликогенот може да се синтетизира во речиси сите органи и ткива. Сепак, најголем крај
Анаеробен гликолиз
Во зависност од функционалната состојба на телото, клетките на органите и ткивата може да бидат во услови на доволно снабдување со кислород и тестирање на нејзиниот недостаток,
Аеробни гликолиз (хексозодифантска патека)
Hexosodiphoshat патот. Ова е класичниот пат на аеробниот катаболизам на јаглени хидрати во ткивата тече во цитоплазмата до фазата на формирање на пируват и завршува во митохондриите со формирањето на con
Хексомонофосфат патека
Хексомонофосфат пат на трансформација на гликоза во ткивата, реакција хемија. Оксидацијата на гликоза на овој пат тече во цитоплазмата на клетките и е претставена со две последователни гранки
Gloundoegenesis.
Глуконеогенеза Главните извори на гликоза за човечкото тело се: 1. Јаглени хидрати; 2. Гликоген ткаенини; 3. Gloungenesis. Слоногенеза е
Главни липиди човечко тело нивната биолошка улога
Липид се нарекува сложени органски супстанции на биолошката природа нерастворлива во вода, но растворлив во органски растворувачи. Липидите се главната храна. Тие П.
Варење на липиди, престој на маснотии
Варење на липиди. Апликациите од храната Липид во усната шуплина се само механичка обработка. Липолитичките ензими во усната шуплина не се формираат. Зголемување на маснотиите
Липопротеини на крв
Според тоа, липидите се нерастворливи во соединенија со вода, поради што се потребни посебни превозници, кои се растворливи во вода. Таквите транспортни форми се липопротеини.
Оксидација на повисоки масни киселини
Масти ткаенина составена од адипозоцити врши специфична улога во липидниот метаболизам. Околу 65% од масата на масното ткиво паѓа врз уделот на триацил глицерол (ознака) во очекување во него - тие замислуваат
Биосинтеза GWK во ткива
Биосинтезата GWK се јавува во ендоплазматската мобилна мрежа. Заменети LGK (сите граници и непредвидени со една двојна врска) се синтетизираат во клетките од ацетил-коа. Услови за Б.
Cholesterol размена
Размена на холестерол. Холестерол е претходник во стероидната синтеза: жолчни киселини, стероидни хормони, витамин Д3ХХОЕРИН е задолжителен структурен
Дигестивни протеини
Варење на протеини во дигестивниот тракт, хранливите протеини се подложени на хидролитичко разделување под дејство на протеолитички ензими (класа - хидролаза, подкласа - пептидаза).
Намалување на амино киселини, неутрализација на гнили производи
На гнили аминокиселински амино киселини кои не беа подложени на вшмукување се запишат во дебелото црево, каде што тие се подложени на гнили. Амино киселината гние е процес на распаѓање на амино киселини
Метаболизам на амино киселини
Метаболизмот на амино киселини од изворите на амино киселини во ќелијата е: 1. протеини од храна по нивната хидролиза во органите за дигестија; 2. Синтеза на заменливи амино киселини;
Начини за неутрализирање на амонијак
Амонијак е формиран од амино киселини за време на распаѓањето на други соединенија кои содржат азот (биогени амини, нуклеотиди). Значителен дел од амонијак се формира во голема црева во ротација. Се апсорбира во ЦР.
Регулирање на метаболизмот
Молекули на сигнал. Главните задачи на регулирање на метаболизмот и мобилните функции: 1. Интрацелуларна и меѓусебна координација на метаболичките процеси; 2. Исклучок "неактивен
Хормони Хипоталамус
Химоталамус хипоталамус Хипоталамус е компонента и посебен "излезен канал" на лимбичкиот систем. Ова е Одделот за средно мозокот контролира различни параметри на ГОМ
Хормони хипофизни жлезди
Хормони хипофизни жлезди
Хормони хипофизни жлезди во хипофизата ја распределуваат фронтот (аденогипид) и задни влогови (неврохипофиза). Аденологипос хормоните може да се поделат на 3 групи во зависност
Биосинтеза Uodthththinthrins.
Синтеза на uodthththnth се јавува во составот на протеини - тироглобулин, кој е во фоликулите на тироидната жлезда. Thyroglobulin е гликопротеин кој содржи 115 тиросински остатоци. Р
LIPID размена
Во црниот дроб на масното ткиво, хормоните ја стимулираат липолизата. Овие ефекти врз размена на јаглени хидрати и липиди се поврзани со зголемување на клеточната чувствителност на адреналинската акција под влијание на колибата на тироидната жлезда
Хидосокција
Во детството, падот на секрецијата води до одложување на физичкиот и менталниот развој (кретинизам). Кај возрасни, тешка манифестација на недостаток на тироидни хормони се меша
Хиперсекреција
Дифузна токсична густа (заболена болест) Најчеста болест придружена со зголемени производи за производство на yodthnin. Во оваа болест, големини на тироидната жлезда се зголемуваат и
Хормони на паратироидни жлезди
Преседанието е синтетизиран во паратироидните жлезди и се состои од 84 аминокиселини остатоци. Хормонот е зачуван во секреторни гранули. ПТГ секрецијата е регулирана со нивото на калциум во крвта: кога
Хормони на генитални жлезди
Хормоните на сексуалните жлезди во хемиска природа се стероиди. Одвојува: 1. Андрогени; 2. Естрогени; 3. Прогестини.
Хормони надбубрежни жлезди
Хормони на надбубрежните надбубрежни жлезди - жлезди на внатрешна секреција, во која се разликува кортикалната и braindatus. Во кортексот слој, хормоните на стероидната природа се синтетизираат, во мозокот
Панкреатски хормони
Панкреатски хормони на функцијата на панкреасот: · Ексокрин; · Ендокрин. Ексокрината функција лежи во синтезата и секрецијата на дигестивната фарма
Испитни прашања
Фармацевтски факултет (одделот за кореспонденција) Испитување Прашања за биолошка хемија за учениците од 3 курсеви (6 семестар) 1. Биохемија, неговите задачи. Биохемија врска со f
